Temperate Fruit Trees Under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Determination of Flavonoids and Hydroxycinnamic Acids in the Herb
The Pharma Innovation Journal 2020; 9(1): 43-46 ISSN (E): 2277- 7695 ISSN (P): 2349-8242 NAAS Rating: 5.03 Determination of flavonoids and hydroxycinnamic TPI 2020; 9(1): 43-46 © 2020 TPI acids in the herb of common agrimony by HPLC www.thepharmajournal.com Received: 24-11-2019 method Accepted: 28-12-2019 NM Huzio NM Huzio, AR Grytsyk, LI Budniak and IR Bekus Department of Pharmacy, Ivano-Frankivsk National Medical University, Ivano- Abstract Frankivsk, 76000, Ukraine The creation of new herbal products and the improvement of their production technologies is an important area of pharmaceutical science. A valuable source of biologically active substances is a AR Grytsyk representative of the Rose family (Rosaceae) common agrimony (Agrimonia eupatoria L.). Remedies on Department of Pharmacy, the basis of common agrimony are used to increase the secretion of the digestive glands, as appetizing, Ivano-Frankivsk National choleretic, hemostatic, astringent, anti-diuretic and anti-inflammatory agent. The chemical composition Medical University, Ivano- of the common agrimony herb grown on the territory of Ukraine is poorly studied, so conducting an in- Frankivsk, 76000, Ukraine depth phytochemical study of the raw material of this type of plant is relevant. The purpose of the work was the determination of individual flavonoids and hydroxycinnamic acids by high performance liquid LI Budniak chromatography (HPLC). The determination of the qualitative composition and the quantitative content Department of Pharmacy Management, Economics and of the individual flavonoids and hydroxycinnamic acids in the common agrimony herb were performed Technology, I. Horbachevsky by HPLC on an Agilent 1200 chromatograph. Ternopil National Medical As a result of the conducted studies, the quantitative content of 4 flavonoids – isoquercitrin (916.73 University, Ternopil, 46000, µg/g), neohesperidin (3850.93 µg/g), naringenin (308.28 µg/g) and luteolin (332.13 µg/g) was found and Ukraine determined in the common agrimony herb. -

2021 Girls Spring Season
2021 GIRLS' SPRING PROGRAM SEASON INFORMATION PACKET LAST UPDATED: WEDNESDAY, APRIL 7TH @ 10:00PM 2020-21 RETURN TO PLAY - MAKING YOUR SAFETY A PRIORITY C R E A T E D B Y V C U N I T E D S T A F F U S I N G R E S T O R E I L L I N O I S A N D J V A / U S A V / A A U V O L L E Y B A L L G U I D E L I N E S 2021 SPRING TRYOUTS 2020-21 Seaon - Return To Play - Making Your Safety Our Priority GET READY FOR THE 2021 SPRING SEASON WHY TRYOUTS? Even though we anticipate that the Pre-TRryouEt Cl-iniTcs aRre a gYreatO way Uto prTepar eC for tLhe uIpNcomiIngC club season early sessions will be in-house leagues, or simply keep your skills sharp during the year. Each session will focus on a we need to accomplish two goals with range of skills and include drills to sharpen your overall game and build our tryouts. First, to create a competitive training environment with your confidence as you prepare for the spring club season. players of similar ability and objectives. Second, is to be in a position to quickly U17 U16 U15 move to teams/tournament play when SATURDAY, APRIL 17 SATURDAY, APRIL 17 SATURDAY, APRIL 17 Illinois determines it is safe to do so. 9A-11A OR 1P-3P 9A-11A OR 1P-3P 9A-11A OR 1P-3P COST: $30 COST: $30 COST: $30 AGE GROUPS USA Volleyball and AAU Volleyball have U14 U13 U12-U11 changed the birthdate cutoff starting SATURDAY, APRIL 17 SATURDAY, APRIL 17 SATURDAY, APRIL 17 with the upcoming season. -

Increasing Day-Length Induces Spring Flushing of Tropical Dry Forest Trees in the Absence of Rain
Trees (2002) 16:445–456 DOI 10.1007/s00468-002-0185-3 ORIGINAL ARTICLE Guillermo Rivera · Stephen Elliott · Linda S. Caldas Guillermo Nicolossi · Vera T. R. Coradin Rolf Borchert Increasing day-length induces spring flushing of tropical dry forest trees in the absence of rain Received: 10 September 2001 / Accepted: 26 March 2002 / Published online: 20 July 2002 © Springer-Verlag 2002 Abstract In many conspecific trees of >50 species high- synthetic gain in tropical forests with a relatively short ly synchronous bud break with low inter-annual varia- growing season. tion was observed during the late dry season, around the spring equinox, in semideciduous tropical forests of Keywords Bud break · Phenology · Photoperiodic Argentina, Costa Rica, Java and Thailand and in tropical control · Tropical semideciduous forests savannas of Central Brazil. Bud break was 6 months out of phase between the northern and southern hemispheres and started about 1 month earlier in the subtropics than Introduction at lower latitudes. These observations indicate that “spring flushing”, i.e., synchronous bud break around the In cold-temperate forests, vegetative phenology of all spring equinox and weeks before the first rains of the broad-leaved trees is strongly synchronized by winter wet season, is induced by an increase in photoperiod of cold. In contrast, severe seasonal drought does not syn- 30 min or less. Spring flushing is common in semidecid- chronize vegetative phenology in tropical semideciduous uous forests characterized by a 4–6 month dry season forests with a dry season of 4–6 months and annual rain- and annual rainfall of 800–1,500 mm, but rare in neo- fall between 800 and 1,500 mm. -
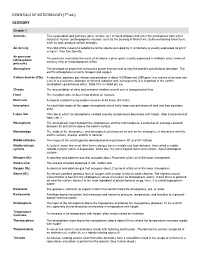
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

INDEX for 2011 HERBALPEDIA Abelmoschus Moschatus—Ambrette Seed Abies Alba—Fir, Silver Abies Balsamea—Fir, Balsam Abies
INDEX FOR 2011 HERBALPEDIA Acer palmatum—Maple, Japanese Acer pensylvanicum- Moosewood Acer rubrum—Maple, Red Abelmoschus moschatus—Ambrette seed Acer saccharinum—Maple, Silver Abies alba—Fir, Silver Acer spicatum—Maple, Mountain Abies balsamea—Fir, Balsam Acer tataricum—Maple, Tatarian Abies cephalonica—Fir, Greek Achillea ageratum—Yarrow, Sweet Abies fraseri—Fir, Fraser Achillea coarctata—Yarrow, Yellow Abies magnifica—Fir, California Red Achillea millefolium--Yarrow Abies mariana – Spruce, Black Achillea erba-rotta moschata—Yarrow, Musk Abies religiosa—Fir, Sacred Achillea moschata—Yarrow, Musk Abies sachalinensis—Fir, Japanese Achillea ptarmica - Sneezewort Abies spectabilis—Fir, Himalayan Achyranthes aspera—Devil’s Horsewhip Abronia fragrans – Sand Verbena Achyranthes bidentata-- Huai Niu Xi Abronia latifolia –Sand Verbena, Yellow Achyrocline satureoides--Macela Abrus precatorius--Jequirity Acinos alpinus – Calamint, Mountain Abutilon indicum----Mallow, Indian Acinos arvensis – Basil Thyme Abutilon trisulcatum- Mallow, Anglestem Aconitum carmichaeli—Monkshood, Azure Indian Aconitum delphinifolium—Monkshood, Acacia aneura--Mulga Larkspur Leaf Acacia arabica—Acacia Bark Aconitum falconeri—Aconite, Indian Acacia armata –Kangaroo Thorn Aconitum heterophyllum—Indian Atees Acacia catechu—Black Catechu Aconitum napellus—Aconite Acacia caven –Roman Cassie Aconitum uncinatum - Monkshood Acacia cornigera--Cockspur Aconitum vulparia - Wolfsbane Acacia dealbata--Mimosa Acorus americanus--Calamus Acacia decurrens—Acacia Bark Acorus calamus--Calamus -

Complete Iowa Plant Species List
!PLANTCO FLORISTIC QUALITY ASSESSMENT TECHNIQUE: IOWA DATABASE This list has been modified from it's origional version which can be found on the following website: http://www.public.iastate.edu/~herbarium/Cofcons.xls IA CofC SCIENTIFIC NAME COMMON NAME PHYSIOGNOMY W Wet 9 Abies balsamea Balsam fir TREE FACW * ABUTILON THEOPHRASTI Buttonweed A-FORB 4 FACU- 4 Acalypha gracilens Slender three-seeded mercury A-FORB 5 UPL 3 Acalypha ostryifolia Three-seeded mercury A-FORB 5 UPL 6 Acalypha rhomboidea Three-seeded mercury A-FORB 3 FACU 0 Acalypha virginica Three-seeded mercury A-FORB 3 FACU * ACER GINNALA Amur maple TREE 5 UPL 0 Acer negundo Box elder TREE -2 FACW- 5 Acer nigrum Black maple TREE 5 UPL * Acer rubrum Red maple TREE 0 FAC 1 Acer saccharinum Silver maple TREE -3 FACW 5 Acer saccharum Sugar maple TREE 3 FACU 10 Acer spicatum Mountain maple TREE FACU* 0 Achillea millefolium lanulosa Western yarrow P-FORB 3 FACU 10 Aconitum noveboracense Northern wild monkshood P-FORB 8 Acorus calamus Sweetflag P-FORB -5 OBL 7 Actaea pachypoda White baneberry P-FORB 5 UPL 7 Actaea rubra Red baneberry P-FORB 5 UPL 7 Adiantum pedatum Northern maidenhair fern FERN 1 FAC- * ADLUMIA FUNGOSA Allegheny vine B-FORB 5 UPL 10 Adoxa moschatellina Moschatel P-FORB 0 FAC * AEGILOPS CYLINDRICA Goat grass A-GRASS 5 UPL 4 Aesculus glabra Ohio buckeye TREE -1 FAC+ * AESCULUS HIPPOCASTANUM Horse chestnut TREE 5 UPL 10 Agalinis aspera Rough false foxglove A-FORB 5 UPL 10 Agalinis gattingeri Round-stemmed false foxglove A-FORB 5 UPL 8 Agalinis paupercula False foxglove -

Ontogenetic Studies on the Determination of the Apical Meristem In
Ontogenetic studies on the determination of the apical meristem in racemose inflorescences D i s s e r t a t i o n Zur Erlangung des Grades Doktor der Naturwissenschaften Am Fachbereich Biologie Der Johannes Gutenberg-Universität Mainz Kester Bull-Hereñu geb. am 19.07.1979 in Santiago Mainz, 2010 CONTENTS SUMMARY OF THE THESIS............................................................................................ 1 ZUSAMMENFASSUNG.................................................................................................. 2 1 GENERAL INTRODUCTION......................................................................................... 3 1.1 Historical treatment of the terminal flower production in inflorescences....... 3 1.2 Structural understanding of the TF................................................................... 4 1.3 Parallel evolution of the character states referring the TF............................... 5 1.4 Matter of the thesis.......................................................................................... 6 2 DEVELOPMENTAL CONDITIONS FOR TERMINAL FLOWER PRODUCTION IN APIOID UMBELLETS...................................................................................................... 7 2.1 Introduction...................................................................................................... 7 2.2 Materials and Methods..................................................................................... 9 2.2.1 Plant material.................................................................................... -

Agrimonia Eupatoria L. and Cynara Cardunculus L. Water Infusions: Phenolic Profile and Comparison of Antioxidant Activities
Article Agrimonia eupatoria L. and Cynara cardunculus L. Water Infusions: Phenolic Profile and Comparison of Antioxidant Activities Anika Kuczmannová 1,†, Peter Gál 1,2,3,4,†,*, Lenka Varinská 2,3, Jakub Treml 5, Ivan Kováˇc 3,6, Martin Novotný 3,7, Tomáš Vasilenko 3,8, Stefano Dall’Acqua 9, Milan Nagy 1 and Pavel Muˇcaji 1,* Received: 29 September 2015 ; Accepted: 4 November 2015 ; Published: 18 November 2015 Academic Editor: Maurizio Battino 1 Department of Pharmacognosy and Botany, Faculty of Pharmacy, Comenius University, Odbojárov 10, 832 32 Bratislava, Slovakia; [email protected] (A.K.); [email protected] (M.N.) 2 Department of Pharmacology, Faculty of Medicine, Pavol Jozef Šafárik University, Trieda SNP 1, 040 11 Košice, Slovakia; [email protected] 3 Department for Biomedical Research, East-Slovak Institute of Cardiovascular Diseases, Inc., Ondavská 8, 040 11 Košice, Slovakia; [email protected] (I.K.); [email protected] (M.N.); [email protected] (T.V.) 4 Institute of Anatomy, 1st Faculty of Medicine, Charles University, U nemocnice 2, 128 00 Prague, Czech Republic 5 Department of Molecular Biology and Pharmaceutical Biotechnology, Faculty of Pharmacy, University of Veterinary and Pharmaceutical Sciences, Palackého 1-3, 612 42 Brno, Czech Republic; [email protected] 6 2nd Department of Surgery, Pavol Jozef Šafárik University and Louise Pasteur University Hospital, 041 90 Košice, Slovakia 7 Department of Infectology and Travel Medicine, Pavol Jozef Šafárik University and Louise Pasteur University Hospital, 041 90 Košice, Slovakia 8 Department of Surgery, Pavol Jozef Šafárik University and Košice-Šaca Hospital, 040 15 Košice-Šaca, Slovakia 9 Department of Pharmaceutical and Pharmacological Sciences, University of Padova, Via F. -

Spring Winter Summer Autumn
• Always drive on good, properly inflated issouri is a state of four seasons tires. and each season has its own unique road conditions. Missouri driving • Know and obey all traffic laws. cannot be categorized entirely into spring, summer, autumn, or winter. Nature some- • Be ready to adjust your speed to be ap- times mixes our four seasons together, and propriate for constantly changing driving this can cause problems when we travel. conditions. This brochure has been prepared to give you some tips on how to handle our Finally, let’s all work together, so fewer many varied driving conditions. people will become traffic crash statistics on Missouri’s highways. Spring Buckle Up Missouri! • Never drive when you have been drink- ing alcoholic beverages. Summer • Never ride with someone who has been drinking. • If medication directions indicate you should not drive after taking it, don’t do Feel free to call the it. Road Condition Report Hotline at: Produced by: Public Information and Education Division • Have a good attitude when you drive. Be Published by: Autumn patient with others. 1-888-275-6636 Missouri State Highway Patrol 1510 East Elm Street • Give driving your full attention. Behind Or, check the Patrol’s Jefferson City, MO 65101 the wheel is no place to read, put on 573-751-3313 makeup, or talk on the cell phone. web site at: V/TDD 573-751-3313 email: [email protected] • How about those eyes? Don’t be vain. If www.mshp.dps.mo.gov www.mshp.dps.mo.gov Winter you need glasses, wear them. -

A Review of Droughts in the African Continent: a Geospatial and Long
Discussion Paper | Discussion Paper | Discussion Paper | Discussion Paper | Open Access Hydrol. Earth Syst. Sci. Discuss., 11, 2679–2718, 2014 Hydrology and www.hydrol-earth-syst-sci-discuss.net/11/2679/2014/ Earth System doi:10.5194/hessd-11-2679-2014 © Author(s) 2014. CC Attribution 3.0 License. Sciences Discussions This discussion paper is/has been under review for the journal Hydrology and Earth System Sciences (HESS). Please refer to the corresponding final paper in HESS if available. A review of droughts in the African continent: a geospatial and long-term perspective I. Masih1, S. Maskey1, F. E. F. Mussá1,2, and P. Trambauer1 1UNESCO-IHE, Institute for Water Education, P.O. Box 3015, 2601 DA Delft, the Netherlands 2Eduardo Mondlane University, Faculty of Engineering, Av. de Moçambique km 1.5, C. Postal 257, Maputo, Mozambique Received: 31 January 2014 – Accepted: 3 February 2014 – Published: 6 March 2014 Correspondence to: I. Masih ([email protected]) Published by Copernicus Publications on behalf of the European Geosciences Union. 2679 Discussion Paper | Discussion Paper | Discussion Paper | Discussion Paper | Abstract This paper presents a comprehensive review and analysis of the available literature and information on droughts to build a continental, regional and country level perspective on geospatial and temporal variation of droughts in Africa. The study is based on 5 the review and analysis of droughts occurred during 1900–2013 as well as evidence available from past centuries based on studies on the lake sediment analysis, tree- ring chronologies and written and oral histories and future predictions from the global climate change models. -

Antioxidative Potential of Agrimonia Eupatoria L
Science & Technologies ANTIOXIDATIVE POTENTIAL OF AGRIMONIA EUPATORIA L. Ivanova D., Tasinov O*., Vankova D., Kiselova-Kaneva Y. Department of Biochemistry, Molecular Medicine and Nutrigenomics, Medical University “Prof. Dr. P. Stoyanov”, 9002 Varna, Bulgaria; *Corresponding author: [email protected] ABSTRACT In folk medicine Agrimonia eupatoria (Agrimony) is well known with its beneficial effects in various diseases. Based on its use, studies on aqueous and aqueous-alcoholic extracts are directed on elucidating of the antioxidant, antidiabetic and anti-inflammatory activity. We have shown that antioxidant activity of Agrimony is associated with high polyphenolic content, and the extracts modified the expression of pro-inflammatory factors and enzymes from glutathione metabolism in cell culture and animal models. Administration of extracts prevented body mass gain and fat accumulation and normalized serum lipid profile in high fructose fed rats. The herb is shown to be a perspective therapeutic especially for treatment of social significant diseases such as diabetes and obesity, accompanied by low-grade inflammation. Keywords: Agrimonia eupatoria, antioxidant, antidiabetic, anti-inflammatory, glutathione Introduction. Agrimonia eupatoria L. is traditionally used in Bulgarian folk medicine to treat various inflammatory diseases. Studies in the recent years suggest that its therapeutic potential is also related to social significant diseases such as type 2 diabetes. However, the mechanisms of its effects are not well understood. Botanical characteristics. The genus Agrimonia are perennial herbaceous flowering plants classified in Division Magnoliophyta, Class: Magnoliopsida, Family: Rosaceae, Genus: Agrimonia. Two species of the plant, Agrimonia eupatoria and Agrimonia procera, are naturally spread in Bulgaria (16). Agrimony (A. eupatoria) is flowering with 5 parts yellow blossoms. -

Wsn 89 (2017) 90-97 Eissn 2392-2192
Available online at www.worldscientificnews.com WSN 89 (2017) 90-97 EISSN 2392-2192 Biologically active compounds in Agrimoni eupatoria L. and their therapeutic effects Mikołaj Kostryco1, Mirosława Chwil2 Department of Botany, University of Life Sciences in Lublin, 15 Akademicka Str., 20-950 Lublin, Poland 1,2E-mail address: [email protected] , [email protected] ABSTRACT Three species from the genus Agrimonia occur in Poland: Agrimonia eupatoria L., Agrimonia pilosa Ledeb., and Agrimonia procera Wallr. Agrimonia eupatoria is a native plant growing commonly in meadow, scrub, and ruderal communities in lowland areas and in lower montane zones. A. eupatoria species have been classified as edible, cosmetic, and medicinal plants. Herba Agrimoniae is a medicinal raw material from A. eupatoria L. It is a source of many biologically active compounds, mainly flavonoids, tannins, proteins, carbohydrates, and vitamins. A. eupatoria herb is characterised by varied contents of groups of biologically active compounds. The A. eupatoria bioactive compounds exhibit antiviral, antibacterial, anti-inflammatory, anti-carcinogenic, and antioxidant activity. This rich source of health-enhancing substances is used in treatment of disorders of the gastrointestinal system and bile ducts as well as inflammatory diseases of the skin. With their antiseptic and anti- inflammatory properties, A. eupatoria infusions are used to alleviate throat diseases and are recommended to occupational voice users. Keywords: common agrimony, active compounds, therapeutic application, Rosaceae 1. INTRODUCTION The family Rosaceae comprises over 300 plant species occurring worldwide, mainly in the northern hemisphere. Its taxa grow in moderate climate forests as understory species, ( Received 19 October 2017; Accepted 10 November 2017; Date of Publication 22 November 2017 ) World Scientific News 89 (2017) 90-97 saltwater and freshwater marshes, and the Arctic tundra.