Lynx Lynx to SCOTLAND
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

After Long-Term Decline, Are Aspen Recovering in Northern Yellowstone? ⇑ Luke E
Forest Ecology and Management 329 (2014) 108–117 Contents lists available at ScienceDirect Forest Ecology and Management journal homepage: www.elsevier.com/locate/foreco After long-term decline, are aspen recovering in northern Yellowstone? ⇑ Luke E. Painter a, , Robert L. Beschta a, Eric J. Larsen b, William J. Ripple a a Department of Forest Ecosystems and Society, Oregon State University, Corvallis, OR 97331, USA b Department of Geography and Geology, University of Wisconsin – Stevens Point, Stevens Point, WI 54481-3897, USA article info abstract Article history: In northern Yellowstone National Park, quaking aspen (Populus tremuloides) stands were dying out in the Received 18 December 2013 late 20th century following decades of intensive browsing by Rocky Mountain elk (Cervus elaphus). In Received in revised form 28 May 2014 1995–1996 gray wolves (Canis lupus) were reintroduced, joining bears (Ursus spp.) and cougars (Puma Accepted 30 May 2014 concolor) to complete the guild of large carnivores that prey on elk. This was followed by a marked decline in elk density and change in elk distribution during the years 1997–2012, due in part to increased pre- dation. We hypothesized that these changes would result in less browsing and an increase in height of Keywords: young aspen. In 2012, we sampled 87 randomly selected stands in northern Yellowstone, and compared Wolves our data to baseline measurements from 1997 and 1998. Browsing rates (the percentage of leaders Elk Browsing effects browsed annually) in 1997–1998 were consistently high, averaging 88%, and only 1% of young aspen Trophic cascade in sample plots were taller than 100 cm; none were taller than 200 cm. -

Wolf Interactions with Non-Prey
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln USGS Northern Prairie Wildlife Research Center US Geological Survey 2003 Wolf Interactions with Non-prey Warren B. Ballard Texas Tech University Ludwig N. Carbyn Canadian Wildlife Service Douglas W. Smith US Park Service Follow this and additional works at: https://digitalcommons.unl.edu/usgsnpwrc Part of the Animal Sciences Commons, Behavior and Ethology Commons, Biodiversity Commons, Environmental Policy Commons, Recreation, Parks and Tourism Administration Commons, and the Terrestrial and Aquatic Ecology Commons Ballard, Warren B.; Carbyn, Ludwig N.; and Smith, Douglas W., "Wolf Interactions with Non-prey" (2003). USGS Northern Prairie Wildlife Research Center. 325. https://digitalcommons.unl.edu/usgsnpwrc/325 This Article is brought to you for free and open access by the US Geological Survey at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in USGS Northern Prairie Wildlife Research Center by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. 10 Wolf Interactions with Non-prey Warren B. Ballard, Ludwig N. Carbyn, and Douglas W. Smith WOLVES SHARE THEIR ENVIRONMENT with many an wolves and non-prey species. The inherent genetic, be imals besides those that they prey on, and the nature of havioral, and morphological flexibility of wolves has the interactions between wolves and these other crea allowed them to adapt to a wide range of habitats and tures varies considerably. Some of these sympatric ani environmental conditions in Europe, Asia, and North mals are fellow canids such as foxes, coyotes, and jackals. America. Therefore, the role of wolves varies consider Others are large carnivores such as bears and cougars. -

The Cost of Migratory Prey: Seasonal Changes in Semi-Domestic Reindeer Distribution Influences Breeding Success of Eurasian Lynx in Northern Norway
The cost of migratory prey: seasonal changes in semi-domestic reindeer distribution influences breeding success of Eurasian lynx in northern Norway Zea Walton1, Jenny Mattisson2, John D. C. Linnell2, Audun Stien3 and John Odden2 1Dept of Forestry and Wilderness Management, Hedmark College, Koppang, Norway 2Norwegian Inst. for Nature Research (NINA), NO-7484 Trondheim, Norway 3Norwegian Inst. for Nature Research (NINA), Fram Centre, Tromsø, Norway Corresponding author: Zea Walton, Dept of Forestry and Wilderness Management, Hedmark College, Koppang, Norway. E-mail: [email protected] Decision date: 31-Aug-2016 This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as doi: [10.1111/oik.03374]. ‘This article is protected by copyright. All rights reserved.’ Accepted Article Accepted (Abstract) Migratory prey is a widespread phenomenon that has implications for predator–prey interactions. By creating large temporal variation in resource availability between seasons it becomes challenging for carnivores to secure a regular year-round supply of food. Some predators may respond by following their migratory prey, however, most predators are sedentary and experience strong seasonal variation in resource availability. Increased predation on alternative prey may dampen such seasonal resource fluctuations, but reduced reproduction rates in predators is a predicted consequence of migratory primary prey behavior that has received little empirical attention. We used data from 23 GPS collared Eurasian lynx Lynx lynx monitored during 2007–2013 in northern Norway, to examine how spatio-temporal variation in the migratory behavior of semi-domestic reindeer Rangifer tarandus influences lynx spatial organization and reproductive success using estimates of seasonal home range overlap and breeding success. -

A Review of Bird of Prey Persecution in Scotland 2001
PERSECUTION A review of bird of prey persecution in Scotland in 2001 Produced by RSPB Scotland A Review of Bird of Prey Persecution in Scotland 2001 28 January 2003 wpo\ab\BOP1\5232 Birds of Prey Report 2001 Contents 1 Preamble 2 2 Recommendations 2 3 Introduction 3 4 Poisoning 4 5 Direct persecution other than poisoning 5 6 Investigation and prosecution 7 Poisoning incidents 7 Incidents other than poisoning 8 7 Discussion of the general nature of persecution offences8 The law 8 Comparative distribution of 2001 and past incidents 9 Published material that indicates likely offenders 10 Prosecutions 11 8 Identifiable trends in persecution 12 9 Conclusions 12 10 Acknowledgements 13 11 Appendices and Maps 13 Appendix A 13 Appendix B 15 Appendix C 15 Appendix D 18 Table 2 18 Map 1 23 Map 2 24 Map 3 25 Map 4 26 1 Birds of Prey Report 2001 1 Preamble The deliberate destruction of Scotland’s birds of prey has been a prominent issue for many decades. The practice of eliminating all the possible predators of game on shooting estates was a routine procedure in the 19th century with little or no regard to the conservation status of the targeted animals. This resulted in national and regional extinctions of a number of predatory birds and other animals. Many of these extirpated species have made significant recoveries in recent years either through natural re-colonisation or through re- introduction by humans. This implies a reduction in killing sufficient to allow these recoveries or facilitate re-introductions. This generally positive trend has not been universal. -

Manual on Uniform Traffic Control Devices (MUTCD) What Is the MUTCD?
National Committee on Uniform Traffic Control Devices Bus/BRT Applications Introduction • I am Steve Andrle from TRB standing in for Randy McCourt, DKS Associates and 2019 ITE International Vice President • I co-manage with Claire Randall15 TRB public transit standing committees. • I want to bring you up to date on planned bus- oriented improvements to the Manual on Uniform Traffic Control Devices (MUTCD) What is the MUTCD? • Manual on Uniform Traffic Control Devices (MUTCD) – Standards for roadway signs, signals, and markings • Authorized in 23 CFR, Part 655: It is an FHWA document. • National Committee on Uniform Traffic Control Devices (NCUTCD) develops content • Sponsored by 19 organizations including ITE, AASHTO, APTA and ATSSA (American Traffic Safety Services Association) Background • Bus rapid transit, busways, and other bus applications have expanded greatly since the last edition of the MUTCD in 2009 • The bus-related sections need to be updated • Much of the available research speaks to proposed systems, not actual experience • The NCUTCD felt it was a good time to survey actual systems to see what has worked, what didn’t work, and to identify gaps. National Survey • The NCUTCD established a task force with APTA and FTA • Working together they issued a survey in April of 2018. I am sure some of you received it. • The results will be released to the NCUTCD on June 20 – effectively now • I cannot give you any details until the NCUTCD releases the findings Survey Questions • Have you participated in design and/or operations of -

Histories of Value Following Deer Populations Through the English Landscape from 1800 to the Present Day
Holly Marriott Webb Histories of Value Following Deer Populations Through the English Landscape from 1800 to the Present Day Master’s thesis in Global Environmental History 1 Abstract Marriott Webb, H. 2019. Histories of Value: Following Deer Populations Through the English Landscape from 1800 to the Present Day. Uppsala, Department of Archaeology and Ancient His- tory. Imagining the English landscape as an assemblage entangling deer and people throughout history, this thesis explores how changes in deer population connect to the ways deer have been valued from 1800 to the present day. Its methods are mixed, its sources are conversations – human voices in the ongoing historical negotiations of the multispecies body politic, the moot of people, animals, plants and things which shapes and orders the landscape assemblage. These conversations include interviews with people whose lives revolve around deer, correspondence with the organisations that hold sway over deer lives, analysis of modern media discourse around deer issues and exchanges with the history books. It finds that a non-linear increase in deer population over the time period has been accompanied by multiple changes in the way deer are valued as part of the English landscape. Ending with a reflection on how this history of value fits in to wider debates about the proper representation of animals, the nature of non-human agency, and trajectories of the Anthropocene, this thesis seeks to open up new ways of exploring questions about human- animal relationships in environmental history. Keywords: Assemblages, Deer, Deer population, England, Hunting, Landscape, Making killable, Moots, Multispecies, Nativist paradigm, Olwig, Pests, Place, Trash Animals, Tsing, United Kingdom, Wildlife management. -

Quaternary Science Reviews 42 (2012) 74E84
Author's personal copy Quaternary Science Reviews 42 (2012) 74e84 Contents lists available at SciVerse ScienceDirect Quaternary Science Reviews journal homepage: www.elsevier.com/locate/quascirev Phylogeographic, ancient DNA, fossil and morphometric analyses reveal ancient and modern introductions of a large mammal: the complex case of red deer (Cervus elaphus) in Ireland Ruth F. Carden a,*, Allan D. McDevitt b, Frank E. Zachos c, Peter C. Woodman d, Peter O’Toole e, Hugh Rose f, Nigel T. Monaghan a, Michael G. Campana g, Daniel G. Bradley h, Ceiridwen J. Edwards h,i a Natural History Division, National Museum of Ireland (NMINH), Merrion Street, Dublin 2, Ireland b School of Biology and Environmental Science, University College Dublin, Belfield, Dublin 4, Ireland c Naturhistorisches Museum Wien, Vienna, Austria d 6 Brighton Villas, University College Cork, Cork, Ireland e National Parks and Wildlife Service, Killarney National Park, County Kerry, Ireland f Trian House, Comrie, Perthshire PH6 2HZ, Scotland, UK g Department of Human Evolutionary Biology, Harvard University, 11 Divinity Avenue, Cambridge 02138, USA h Molecular Population Genetics Laboratory, Smurfit Institute, Trinity College Dublin, Dublin 2, Ireland i Research Laboratory for Archaeology, University of Oxford, Dyson Perrins Building, South Parks Road, Oxford OX1 3QY, UK article info abstract Article history: The problem of how and when the island of Ireland attained its contemporary fauna has remained a key Received 29 November 2011 question in understanding Quaternary faunal assemblages. We assessed the complex history and origins Received in revised form of the red deer (Cervus elaphus) in Ireland using a multi-disciplinary approach. Mitochondrial sequences of 20 February 2012 contemporary and ancient red deer (dating from c 30,000 to 1700 cal. -

Inverness Local Plan Public Local Inquiry Report- Volume 3
TOWN AND COUNTRY PLANNING (SCOTLAND) ACT 1997 REPORT OF PUBLIC LOCAL INQUIRY INTO OBJECTIONS TO THE INVERNESS LOCAL PLAN VOLUME 3 THE HINTERLAND AND THE RURAL DEVELOPMENT AREA Reporter: Janet M McNair MA(Hons) MPhil MRTPI File reference: IQD/2/270/7 Dates of the Inquiry: 14 April 2004 to 20 July 2004 CONTENTS VOLUME 3 Abbreviations The A96 Corridor Chapter 24 Land north and east of Balloch 24.1 Land between Balloch and Balmachree 24.2 Land at Lower Cullernie Farm Chapter 25 Inverness Airport and Dalcross Industrial Estate 25.1 Inverness Airport Economic Development Initiative 25.2 Airport Safeguarding 25.3 Extension to Dalcross Industrial Estate Chapter 26 Former fabrication yard at Ardersier Chapter 27 Morayhill Chapter 28 Lochside The Hinterland Chapter 29 Housing in the Countryside in the Hinterland 29.1 Background and context 29.2 objections to the local plan’s approach to individual and dispersed houses in the countryside in the Hinterland Objections relating to locations listed in Policy 6:1 29.3 Upper Myrtlefield 29.4 Cabrich 29.5 Easter Clunes 29.6 Culburnie 29.7 Ardendrain 29.8 Balnafoich 29.9 Daviot East 29.10 Leanach 29.11 Lentran House 29.12 Nairnside 29.13 Scaniport Objections relating to locations not listed in Policy 6.1 29.14 Blackpark Farm 29.15 Beauly Barnyards 29.16 Achmony, Balchraggan, Balmacaan, Bunloit, Drumbuie and Strone Chapter 30 Objections Regarding Settlement Expansion Rate in the Hinterland Chapter 31 Local centres in the Hinterland 31.1 Beauly 31.2 Drumnadrochit Chapter 32 Key Villages in the Hinterland -

Wildlife Research Reports
MAMMALS - JULY 2005 WILDLIFE RESEARCH REPORTS JULY 2004 – JUNE 2005 MAMMALS PROGRAM COLORADO DIVISION OF WILDLIFE Research Center, 317 W. Prospect, Fort Collins, CO 80526 The Wildlife Reports contained herein represent preliminary analyses and are subject to change. For this reason, information MAY NOT BE PUBLISHED OR QUOTED without permission of the Author. STATE OF COLORADO Bill Owens, Governor DEPARTMENT OF NATURAL RESOURCES Russell George, Executive Director WILDLIFE COMMISSION Jeffrey Crawford, Chair …………………………………………………………………….…..… Denver Tom Burke, Vice Chair ………………………………….…………...………….…........…Grand Junction Ken Torres, Secretary ……………………………………...…………….……………..……….... Weston Robert Bray………………………………………………….......................................................…Redvale Rick Enstrom………………………………………………………………….………….……...Lakewood Philip James …………………………………………………………………..….………….…Fort Collins Claire M. O’Neal………………………………………………..…………….………..…………..Holyoke Richard Ray ………………………………………………………………………………...Pagosa Springs Robert T. Shoemaker…………………………………………………………….………..…….Canon City Don Ament, Dept. of Ag, Ex-officio…………………………………………………….…….....Lakewood Russell George, Executive Director, Ex-officio……………………………………………..………Denver DIRECTOR’S STAFF Bruce McCloskey, Director Mark Konishi, Deputy Director-Education and Public Affairs Steve Cassin, Chief Financial Officer Jeff Ver Steeg, Assistant Director-Wildlife Programs John Bredehoft, Assistant Director-Field Operations Marilyn Salazar, Assistant Director-Support Services MAMMALS RESEARCH STAFF David Freddy, -
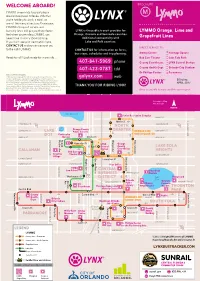
Sunrail.Com Not to Scale
WELCOME ABOARD! BROCHURE LYMMO is your ride to great places M around Downtown Orlando. Whether you’re heading to work, a meal, or one of the many attractions Downtown, LYMMO’s frequent service and bus-only lanes will get you there faster. LYNX is the public transit provider for LYMMO Orange, Lime and And when you’re riding LYMMO, you Orange, Osceola and Seminole counties. never have to worry about parking. Additional connectivity with Grapefruit Lines If you don’t see your destination here, Lake and Polk counties. CONTACT US and we can connect you DIRECT SERVICE TO: to the right LYMMO. CONTACT US for information on fares, bus stops, schedules and trip planning: Amway Center Heritage Square Ready to roll? Look inside for more info... Bob Carr Theater Lake Eola Park 407-841-5969 phone County Courthouse LYNX Central Station 407-423-0787 tdd County Health Dept Orlando City Stadium Dr Phillips Center Parramore Notice of Title VI Rights: LYNX operates its programs and services without regard to race, color, golynx.com web religion, gender, age, national origin, disability, or family status in accordance with Title VI of the Civil Rights Act. Any person who believes Effective: he or she has been aggrieved by any unlawful discriminatory practice APRIL 2017 related to Title VI may file a complaint in writing to LYNX Title VI Officer Desna Hunte, 455 N. Garland Avenue, Orlando, Florida 32801 or by calling THANK YOU FOR RIDING LYNX! 407-254-6117, email [email protected] or www.golynx.com. Information in other languages or accessible formats available upon request. -

Ecology of Red Deer a Research Review Relevant to Their Management in Scotland
Ecologyof RedDeer A researchreview relevant to theirmanagement in Scotland Instituteof TerrestrialEcology Natural EnvironmentResearch Council á á á á á Natural Environment Research Council Institute of Terrestrial Ecology Ecology of Red Deer A research review relevant to their management in Scotland Brian Mitchell, Brian W. Staines and David Welch Institute of Terrestrial Ecology Banchory iv Printed in England by Graphic Art (Cambridge) Ltd. ©Copyright 1977 Published in 1977 by Institute of Terrestrial Ecology 68 Hills Road Cambridge CB2 11LA ISBN 0 904282 090 Authors' address: Institute of Terrestrial Ecology Hill of Brathens Glassel, Banchory Kincardineshire AB3 4BY Telephone 033 02 3434. The Institute of Terrestrial Ecology (ITE) was established in 1973, from the former Nature Conservancy's research stations and staff, joined later by the Institute of Tree Biology and the Culture Centre of Algae and Protozoa. ITE contributes to and draws upon the collective knowledge of the fourteen sister institutes which make up the Natural Environment Research Council, spanning all the environmental sciences. The Institute studies the factors determining the structure, composition and processes of land and freshwater systems, and of individual plant and animal species. It is developing a Sounder scientific basis for predicting and modelling environmental trends arising from natural or man-made change. The results of this research are available to those responsible for the protection, management and wise use of our natural resources. Nearly half of ITE'Swork is research commissioned by customers, such as the Nature Conservancy Council who require information for wildlife conservation, the Forestry Commission and the Department of the Environment. The remainder is fundamental research supported by NERC. -

Information and Analysis Bulletin on Animal Poaching and Smuggling N°6 / 1St July - 30Th Septembre 2014
Information and analysis bulletin on animal poaching and smuggling n°6 / 1st July - 30th Septembre 2014 Contents Introduction Insects and Arachnids 3 Seahorses Sea 4 Cruelty to animals is pouring and flooding on Tridacna and Queen Conches 5 each and every one of the 134 pages of ”On the Sea Cucumbers 5 Trail ” n°6. Rainbows are rare. And for that are all Fishes 6 the more beautiful. Marine Mammals 8 Opening of a delphinium in Pattaya and a Several recent studies suggest that violence on projected opening in Phuket 8 animals and abuse of animal’s weakness show Marine Turtles 9 predisposition to violence against humans and that witnessing domestic violence or being Tortoises and Freshwater Turtles 13 submitted to parental harassment leads to a Snakes 20 predisposition to violence against animals. Sauria 23 Violence is a viral and vicious circle. Crocodilians 23 Multi-Species Reptiles 24 The FBI in the United States of America now considers that mistreatment, individual Amphibia 25 violence, organized abuse and torture against animals are crimes against society. To start Birds 26 in 2016, a database listing animal sadists The new European commissioner for should help - that is the purpose of the FBI’s the environment 32 new approach - to predict criminality against humans. Pangolins 35 Writers and great thinkers in the ancient Primates 40 centuries had predicted that practice or Ebola 42 witnessing of cruelty inflicted on animals would lead to barbaric treatment towards and among Felines 49 humans. ” After people in Rome were tamed Released on bail without caution 53 to the spectacle of murdering animals, it came the turn of men and the gladiators.