Gravel Bar Inundation Frequency: an Important Parameter for Understanding Riparian Corridor Dynamics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lower Wisconsin River Main Stem
LOWER WISCONSIN RIVER MAIN STEM The Wisconsin River begins at Lac Vieux Desert, a lake in Vilas County that lies on the border of Wisconsin and the Lower Wisconsin River Upper Pennisula in Michigan. The river is approximately At A Glance 430 miles long and collects water from 12,280 square miles. As a result of glaciation across the state, the river Drainage Area: 4,940 sq. miles traverses a variety of different geologic and topographic Total Stream Miles: 165 miles settings. The section of the river known as the Lower Wisconsin River crosses over several of these different Major Public Land: geologic settings. From the Castle Rock Flowage, the river ♦ Units of the Lower Wisconsin flows through the flat Central Sand Plain that is thought to State Riverway be a legacy of Glacial Lake Wisconsin. Downstream from ♦ Tower Hill, Rocky Arbor, and Wisconsin Dells, the river flows through glacial drift until Wyalusing State Parks it enters the Driftless Area and eventually flows into the ♦ Wildlife areas and other Mississippi River (Map 1, Chapter Three ). recreation areas adjacent to river Overall, the Lower Wisconsin River portion of the Concerns and Issues: Wisconsin River extends approximately 165 miles from the ♦ Nonpoint source pollution Castle Rock Flowage dam downstream to its confluence ♦ Impoundments with the Mississippi River near Prairie du Chien. There are ♦ Atrazine two major hydropower dams operate on the Lower ♦ Fish consumption advisories Wisconsin, one at Wisconsin Dells and one at Prairie Du for PCB’s and mercury Sac. The Wisconsin Dells dam creates Kilbourn Flowage. ♦ Badger Army Ammunition The dam at Prairie Du Sac creates Lake Wisconsin. -

Watershed Technical Analysis
Trout Creek Watershed Stormwater Management Plan SECTION III: WATERSHED TECHNICAL ANALYSIS A. Watershed Modeling An initial step this study of the Trout Creek watershed was the selection of a stormwater simulation model to be used in the analysis. To provide a reasonable estimate of existing flows within the watershed to serve as a baseline for the study of stormwater management and to effectively evaluate the hydrologic response of the watershed to various changes in stormwater management it was necessary to select a model which: • Modeled design storms of various durations and frequencies to produce routed hydrographs which could be combined • Was adaptable to the size of subwatersheds in this study • Could evaluate specific physical characteristics of the rainfall-runoff process • Did not require an excessive amount of input data yet yielded reliable results The model selected for use in this study was the U. S. Army Corps of Engineers, Hydrologic Engineering Center, Hydrologic Modeling System (HEC-HMS) for the following reasons: • It had been developed at the Hydrologic Engineering Center specifically for the analysis of the timing of surface flow contributions to peak rates at various locations in a watershed • Although originally developed as an urban runoff simulation model, data requirements make it easily adaptable to a rural situation • Input parameters provide a flexible calibration process • It has the ability to analyze reservoir or detention basin routing effects • It is accepted by the Pennsylvania Department of Environmental Protection Although other models, such as TR-20, may provide essentially the same results as the HEC-HMS, HMS’s ability to compare subwatershed contributions in a peak flow presentation table make it specifically attractive for this study. -
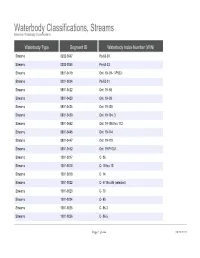
Waterbody Classifications, Streams Based on Waterbody Classifications
Waterbody Classifications, Streams Based on Waterbody Classifications Waterbody Type Segment ID Waterbody Index Number (WIN) Streams 0202-0047 Pa-63-30 Streams 0202-0048 Pa-63-33 Streams 0801-0419 Ont 19- 94- 1-P922- Streams 0201-0034 Pa-53-21 Streams 0801-0422 Ont 19- 98 Streams 0801-0423 Ont 19- 99 Streams 0801-0424 Ont 19-103 Streams 0801-0429 Ont 19-104- 3 Streams 0801-0442 Ont 19-105 thru 112 Streams 0801-0445 Ont 19-114 Streams 0801-0447 Ont 19-119 Streams 0801-0452 Ont 19-P1007- Streams 1001-0017 C- 86 Streams 1001-0018 C- 5 thru 13 Streams 1001-0019 C- 14 Streams 1001-0022 C- 57 thru 95 (selected) Streams 1001-0023 C- 73 Streams 1001-0024 C- 80 Streams 1001-0025 C- 86-3 Streams 1001-0026 C- 86-5 Page 1 of 464 09/28/2021 Waterbody Classifications, Streams Based on Waterbody Classifications Name Description Clear Creek and tribs entire stream and tribs Mud Creek and tribs entire stream and tribs Tribs to Long Lake total length of all tribs to lake Little Valley Creek, Upper, and tribs stream and tribs, above Elkdale Kents Creek and tribs entire stream and tribs Crystal Creek, Upper, and tribs stream and tribs, above Forestport Alder Creek and tribs entire stream and tribs Bear Creek and tribs entire stream and tribs Minor Tribs to Kayuta Lake total length of select tribs to the lake Little Black Creek, Upper, and tribs stream and tribs, above Wheelertown Twin Lakes Stream and tribs entire stream and tribs Tribs to North Lake total length of all tribs to lake Mill Brook and minor tribs entire stream and selected tribs Riley Brook -

Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington
Western Washington University Western CEDAR WWU Graduate School Collection WWU Graduate and Undergraduate Scholarship Winter 1993 Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington Roger G. Bertschi Western Washington University, [email protected] Follow this and additional works at: https://cedar.wwu.edu/wwuet Part of the Geology Commons Recommended Citation Bertschi, Roger G., "Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington" (1993). WWU Graduate School Collection. 717. https://cedar.wwu.edu/wwuet/717 This Masters Thesis is brought to you for free and open access by the WWU Graduate and Undergraduate Scholarship at Western CEDAR. It has been accepted for inclusion in WWU Graduate School Collection by an authorized administrator of Western CEDAR. For more information, please contact [email protected]. CHANNEL CHANGES AND FLOOD FREQUENCY ON THE UPPER MAIN STEM OF THE NOOKSACK RIVER, WHATCOM COUNTY, WASHINGTON by Roger G. Bertschi Accepted in Partial Completion of the Requirements for the Degree Master of Science Advisory Committee MASTER'S THESIS In presenting this thesis in partial fulfillment of the requirements for a master's degree at Western Washington University, I agree that the Library shall make its copies freely available for inspection. I further agree that extensive copying of this thesis is allowable only for scholarly purposes. It is understood, however, that any copying or publication of this thesis for commercial purposes, or for financial gain, shall not be allowed without mv permission. Signature Date MASTER’S THESIS In presenting this thesis in partial fulfillment of the requirements for a master’s degree at Western Washington University, I grant to Western Washington University the non-exclusive royalty-free right to archive, reproduce, distribute, and display the thesis in any and all forms, including electronic format, via any digital library mechanisms maintained by WWU. -

Memorandum 6E. Review of Main Stem Restoration Project; Golden
Memorandum To: Bassett Creek Watershed Management Commission From: Barr Engineering Co. Subject: Item 6E – Review of Main Stem Restoration Project; Golden Valley Rd. to Irving Ave. N. – 50% Development Plans (CIP 2012 CR) BCWMC September 19, 2013 Meeting Agenda Date: September 12, 2013 Project: 23270051 2013 626 6E. Review of Main Stem Restoration Project; Golden Valley Rd. to Irving Ave. N. – 50% Development Plans (CIP 2012 CR) Summary Proposed Work: Main Stem of Bassett Creek Restoration Project (CIP 2012 CR) Basis for Commission Review: 50% plan review Change in Impervious Surface: N.A. Recommendation: Conditional Approval The Minneapolis Park and Recreation Board (MPRB) Main Stem of Bassett Creek restoration project (CIP 2012 CR) is being funded by the BCWMC’s ad valorem levy (via Hennepin County) and by a Minnesota Board of Water and Soil Resources Clean Water Fund Grant. The MPRB provided the 50% design plans to the BCWMC for review and comment, as set forth in the BCWMC CIP project flow chart developed by the TAC. Feasibility Study Summary The Feasibility Report for the 2012 Bassett Creek Main Stem Restoration Project – Golden Valley Road to Irving Avenue North (Barr, June 2011) was completed by the BCWMC to develop approaches to stabilize eroding stream banks along the Main Stem of Bassett Creek. Between Golden Valley Road and Irving Avenue North, Bassett Creek flows through Golden Valley and Minneapolis, and is nearly entirely contained within MPRB-owned land in Theodore Wirth Regional Park, Theodore Wirth Golf Course, and city parks. The goal of the project is to reduce the phosphorus loading to the Main Stem of Bassett Creek by 60 pounds per year and to consolidate sediments in an in-stream pond upstream of Highway 55. -

Surface Water Types and Sediment Distribution Patterns at the Confluence of Mega Rivers: the Solimões-Amazon and Negro Rivers Junction
Surface water types and sediment distribution patterns at the confluence of mega rivers: the Solimões-Amazon and Negro rivers junction Edward Park1, Edgardo M. Latrubesse1 1University of Texas at Austin, Department of Geography and the Environment, Austin, TX, USA Correspondence to: Edward Park, Tel +1-512-230-4603 Fax +1-512-471-5049 University of Texas at Austin, SAC 4.178, 2201 Speedway, Austin, TX 78712, USA Email address: [email protected] This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an ‘Accepted Article’, doi: 10.1002/2014WR016757 This article is protected by copyright. All rights reserved. Abstract Large river channel confluences are recognized as critical fluvial features because both intensive and extensive hydrophysical and geoecological processes take place at this interface. However, identifications of suspended sediment routing patterns through channel junctions and the roles of tributaries on downstream sediment transport in large rivers are still poorly explored. In this paper, we propose a remote sensing-based approach to characterize the spatiotemporal patterns of the post-confluence suspended sediment transport by mapping the surface water distribution in the ultimate example of large river confluence on Earth where distinct water types meet: The Solimões-Amazon (white water) and Negro (black water) rivers. The surface water types distribution was modeled for three different years: average hydrological condition (2007) and two years when extreme events occurred (drought-2005 and flood-2009). -

Tributary Connectivity, Confluence Aggradation and Network Biodiversity
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Loughborough University Institutional Repository Tributary connectivity, confluence aggradation and network biodiversity Stephen P Rice Department of Geography, Loughborough University, Loughborough, LE11 3TU, UK ([email protected]) Accepted GEOMORPHOLOGY March 2016 LUPIN repository version March 2016 1 ABSTRACT In fluvial networks, some confluences are associated with tributary-driven aggradation where coarse sediment is stored, downstream sediment connectivity is interrupted and substantial hydraulic and morphological heterogeneity is generated. To the extent that biological diversity is supported by physical diversity, it has been proposed that the distribution and frequency of tributary-driven aggradation is important for the magnitude and spatial structure of river biodiversity. Relevant ideas are formulated within the Link Discontinuity Concept and the Network Dynamics Hypothesis, but many of the issues raised by these conceptual models have not been systematically evaluated. This paper first tests an automated method for predicting the likelihood of tributary-driven aggradation in three large drainage networks in the Rocky Mountain foothills, Canada. The method correctly identified approximately 75% of significant tributary confluences and 97% of insignificant confluences. The method is then used to evaluate two hypotheses of the Network Dynamics Hypothesis: that linear-shaped basins are more likely to show a longitudinal, downstream decline in tributary-driven aggradation; and that larger and more compact basins contain more confluences with a high probability of impact. The use of a predictive model that included a measure of tributary basin sediment delivery, rather than symmetry ratio alone, mediated the outcomes somewhat, but as anticipated, the number of significant confluences increased with basin size and basin shape was a strong control of the number and distribution of significant confluences. -

Comparison of Methods for Estimating Stream Channel Gradient Using GIS
Comparison of Methods for Estimating Stream Channel Gradient Using GIS David Nagel, John Buffington, and Daniel Isaak USDA Forest Service, Rocky Mountain Research Station Boise Aquatic Sciences Lab Boise, ID September 14, 2006 Special thanks to Sharon Parkes…. StreamStream ChannelChannel GradientGradient Rate of elevation change High Low ComputingComputing GradientGradient Rise / Run = Slope 2115 m 5 m 2110 m 100 m 5 / 100 = .05 = 5% slope ReasonsReasons forfor ModelingModeling StreamStream GradientGradient • Predictor of channel morphology Pool-riffle Plain-bed Step-pool 1% 3 - 4% 10% ReasonsReasons forfor ModelingModeling StreamStream GradientGradient • Estimate distribution of aquatic organisms “Channel gradient and channel morphology appeared to account for the observed differences in salmonid abundance, which reflected the known preference of juvenile coho salmon Oncorhynchus kisutch for pools.” - Hicks, Brendan J. and James D. Hall, 2003 ReasonsReasons forfor ModelingModeling StreamStream GradientGradient • Predict debris flow transport and deposition “Transportation and deposition of material in confined channels are governed primarily by water content of debris, channel gradient, and channel width.” - Fannin, R. J and T. P. Rollerson, 1993 OurOur PurposePurpose forfor ModelingModeling StreamStream GradientGradient • Estimate stream bed grain size to identify salmon spawning habitat 1−n n 1000ρghS 1000cAρ f Sb aA Grain= Size = ( ) ( ) ρ()s − ρg * τ ρ()s − ρk S = channel slope Grain size 16 – 51 mm StudyStudy AreaArea 10,000 km -

Biological Assessment Fact Sheet – Proposed Impaired Water Body
Biological and Aquatic Life Use Attainment Assessment Potash Brook November 15,2005 photo: Potash Brook Site 1.0 prepared by Vermont Department of Environmental Conservation Water Quality Division Biomonitoring and Aquatic Studies Section RA La Rosa Environmental Laboratory 103 South Main Street Waterbury, VT 05671 1 Biological Assessment Summary Fact Sheet – Potash Brook 1. Description of impaired water body: Potash Brook is listed under Vermont’s 2004 303(d) List of Waters, Part A – Impaired Surface Waters in Need of TMDL, as non-support of Aquatic Life Uses (ALUs) with poor to fair biological condition from the mouth to river mile 5.2 and likely including its headwaters. Stormwater is identified as the primary pollutant with problems related to stormwater runoff, land development and soil erosion. The Brook drains much of the town of South Burlington, from its mouth in Lake Champlain in Queen City Park, to just south of Burlington International Airport to 1.5 miles south of Interstate 89. Much its drainage is urban with a short natural wetland between Spear and Farrell Streets. 2. Description of Data used to characterize impairment. Biological data was collected on Potash Brook by the VTDEC from 1987 to 2004, and at several sites by Pioneer Environmental Associates (PEA) for South Burlington in 2001 and 2004. The biological data collected by South Burlington has been approved by VTDEC pursuant to a QA/QC plan, and verification of data quality through sampling and analytical replication between VTDEC and PEA. Macroinvertebrates were sampled 4 times at RM 0.7; 9 times at RM 1.0; 3 times at RM 1.8; twice at RM 1.9; twice at RM 3.0 and 3 times at RM 4.3. -

Spatial Identification of Tributary Impacts in River Networks
, 9 Spatial identification of tributary impacts in river networks 1 2 Christian E. Torgersen , Robert E. Gresswell , Douglas S. Bateman 3 and Kelly M. Burnett4 1 US Geological Survey, Forest and Rangeland Ecosystem Science Center, Cascadia Field Station, University of Washington, College of Forest Resources, USA 2 US Geological Survey, Northern Rocky Mountain Science Center, USA 3 Department of Forest Sciences, Oregon State University, USA 4 US Department of Agriculture, Forest Service, Pacific Northwest Research Station, USA Introduction The ability to assess spatial patterns of ecological conditions in river networks has been confounded by difficulties of measuring and perceiving features that are essentially invisible to observers on land and to aircraft and satellites from above. The nature of flowing water, which is opaque or at best semi -transparent, makes it difficult to visualize fine-scale patterns in habitat and biota at dose range, and the linear topology of river networks complicates the process of scaling up to detect coarse-scale patterns. This spatially incomplete perspective limits our understanding of lotic systems because the scaled character of biotic and abiotic patterns produces different results depending on the method of data collection (Fausch et al., 2002; Hildrew and Giller, 1994). River Confluences, Tributaries and the Fluvial Network Edited by Stephen P. Rice, Andre G. Roy and Bruce L. Rhoads Published 2008 by John Wiley & Sons, Ltd This chapter is a US Government work and is in the public domain in the United States of America. 160 CH 9 SPATIAL IDENTIFICATION OF TRIBUTARY IMPACTS IN RIVER NETWORKS Recent changes in the way river scientists collect data are now filling in these gaps to reveal patterns that raise new questions about the structure and function of river ine mosaics and networks. -

Exercise 4. Watershed and Stream Network Delineation GIS in Water Resources, Fall 2014 Prepared by David G Tarboton and David R
Exercise 4. Watershed and Stream Network Delineation GIS in Water Resources, Fall 2014 Prepared by David G Tarboton and David R. Maidment Purpose The purpose of this exercise is to illustrate watershed and stream network delineation based on digital elevation models using the Hydrology tools in ArcGIS and online services for Hydrology and Hydrologic data. In this exercise, you will select a stream gage location and use online tools to delineate the watershed draining to the gage. National Hydrography and Digital Elevation Model data will be retrieved for this area (Logan River Basin) from online services. You will then perform drainage analysis on a terrain model for this area. The Hydrology tools are used to derive several data sets that collectively describe the drainage patterns of the basin. Geoprocessing analysis is performed to fill sinks and generate data on flow direction, flow accumulation, streams, stream segments, and watersheds. These data are then used to develop a vector representation of catchments and drainage lines from selected points that can then be used in network analysis. This exercise shows how detailed information on the connectivity of the landscape and watersheds can be developed starting from raw digital elevation data, and that this enriched information can be used to compute watershed attributes commonly used in hydrologic and water resources analyses. Learning objectives Do an online watershed delineation and then extract the data for that watershed to perform a more detailed analysis. Identify and properly execute the sequence of Hydrology tools required to delineate streams, catchments and watersheds from a DEM. Evaluate and interpret drainage area, stream length and stream order properties from Terrain Analysis results. -

Memorandum 6A. Review of Main Stem Restoration Project; Golden
Memorandum To: Bassett Creek Watershed Management Commission From: Barr Engineering Co. Subject: Item 6A – Review of Main Stem Restoration Project; Golden Valley Rd. to Irving Ave. N. – 90% Development Plans (CIP 2012 CR) BCWMC March 20, 2014 Meeting Agenda Date: March 12, 2014 Project: 23270051 2014 626 6A. Review of Main Stem Restoration Project; Golden Valley Rd. to Irving Ave. N. – 90% Development Plans (CIP 2012 CR) Summary Proposed Work: Main Stem of Bassett Creek Restoration Project (CIP 2012 CR) Basis for Commission Review: 90% plan review Change in Impervious Surface: N.A. Recommendation: Conditional Approval The Minneapolis Park and Recreation Board (MPRB) Main Stem of Bassett Creek restoration project (CIP 2012 CR) is being funded by the BCWMC’s ad valorem levy (via Hennepin County) and by a Minnesota Board of Water and Soil Resources Clean Water Fund Grant. The MPRB provided the 90% design plans to the BCWMC for review and comment, as set forth in the BCWMC CIP project flow chart developed by the TAC. Feasibility Study Summary The Feasibility Report for the 2012 Bassett Creek Main Stem Restoration Project – Golden Valley Road to Irving Avenue North (Barr, June 2011) was completed by the BCWMC to develop approaches to stabilize eroding stream banks along the Main Stem of Bassett Creek. Between Golden Valley Road and Irving Avenue North, Bassett Creek flows through Golden Valley and Minneapolis, and is nearly entirely contained within MPRB-owned land in Theodore Wirth Regional Park, Theodore Wirth Golf Course, and city parks. The goal of the project is to reduce the phosphorus loading to the Main Stem of Bassett Creek by 60 pounds per year and to consolidate sediments in an in-stream pond upstream of Highway 55.