Surface Water Types and Sediment Distribution Patterns at the Confluence of Mega Rivers: the Solimões-Amazon and Negro Rivers Junction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lower Wisconsin River Main Stem
LOWER WISCONSIN RIVER MAIN STEM The Wisconsin River begins at Lac Vieux Desert, a lake in Vilas County that lies on the border of Wisconsin and the Lower Wisconsin River Upper Pennisula in Michigan. The river is approximately At A Glance 430 miles long and collects water from 12,280 square miles. As a result of glaciation across the state, the river Drainage Area: 4,940 sq. miles traverses a variety of different geologic and topographic Total Stream Miles: 165 miles settings. The section of the river known as the Lower Wisconsin River crosses over several of these different Major Public Land: geologic settings. From the Castle Rock Flowage, the river ♦ Units of the Lower Wisconsin flows through the flat Central Sand Plain that is thought to State Riverway be a legacy of Glacial Lake Wisconsin. Downstream from ♦ Tower Hill, Rocky Arbor, and Wisconsin Dells, the river flows through glacial drift until Wyalusing State Parks it enters the Driftless Area and eventually flows into the ♦ Wildlife areas and other Mississippi River (Map 1, Chapter Three ). recreation areas adjacent to river Overall, the Lower Wisconsin River portion of the Concerns and Issues: Wisconsin River extends approximately 165 miles from the ♦ Nonpoint source pollution Castle Rock Flowage dam downstream to its confluence ♦ Impoundments with the Mississippi River near Prairie du Chien. There are ♦ Atrazine two major hydropower dams operate on the Lower ♦ Fish consumption advisories Wisconsin, one at Wisconsin Dells and one at Prairie Du for PCB’s and mercury Sac. The Wisconsin Dells dam creates Kilbourn Flowage. ♦ Badger Army Ammunition The dam at Prairie Du Sac creates Lake Wisconsin. -

Ethnobotany of Riverine Populations from the Rio Negro, Amazonia (Brazil)
Journal of Ethnobiology 27(1): 46–72 Spring/Summer 2007 ETHNOBOTANY OF RIVERINE POPULATIONS FROM THE RIO NEGRO, AMAZONIA (BRAZIL) ANDRE´ A LEME SILVA,a JORGE TAMASHIROb and ALPINA BEGOSSIc aDepartamento de Ecologia, Instituto de Biocieˆncias Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil, CEP 05580-900 ^[email protected]& bDepartamento de Botaˆnica, UNICAMP Campinas, Sa˜o Paulo, Brazil ^[email protected]& cFisheries and Food Institute, Rua Coronel Quirino 1636, Campinas, Sa˜o Paulo 13025-002, Brazil, and Fisheries Management and Training Program, PREAC-UNICAMP ^[email protected]& ABSTRACT.—This paper presents a comparative study of plant knowledge and use in rural and urban areas in the municipality of Barcelos in the Rio Negro, Amazonas, Brazil, based on a total of 81 interviews. Using diversity indices (Shannon-Wiener), plant knowledge is compared among communities (urban- rural population), and between sex (male-female) and age (older or younger than 40 years) categories within each community. Among our informants, we found quantitative differences concerning the knowledge of medicinal plants between sex and age categories. Some individuals play a key role relating to medicinal plant knowledge, and steps should be taken in order to include them in management and conservation plans. Key words: ethnobotany, diversity indices, plant knowledge and use, Rio Negro, Brazilian Amazon. RESUMO.—Com base em um total de 81 entrevistas, no´s apresentamos um estudo etnobotaˆnico comparativo entre populac¸o˜es urbanas e rurais na municipalidade de Barcelos no Rio Negro, Amazonas, Brasil. Usando´ ındices de diversidade (Shannon-Wiener), o conhecimento de plantas e´ comparado entre as comunidades estudadas (populac¸a˜o urbana e rural), geˆnero (masculino e feminino) e categorias de idade (menos que 40 anos e mais que 40 anos de idade). -

Amazon River Voyage
AMAZON RIVER VOYAGE Over the 38 years that International Expeditions has been leading Amazon River cruises, our guests have enjoyed unrivaled access to the Peruvian rainforest’s most pristine areas and the company of the river’s most knowledgeable guides. Your exciting daily excursions include birding at dawn, school visits in local villages and even piranha fishing! Join IE and be transported to the Amazon of your imagination to discover a rainforest that pulsates with an unrivaled diversity of wildlife. Authentic Amazon River Experience Aboard this luxury Amazon River cruise, you’ll create a lifetime of memories: the joy in the eyes of a village child when you visit their school; the enveloping darkness of the jungle; or the awe possible. of floating along narrow streams accompanied by pink dolphins. Drawing on insight from our native Amazon travel guides, Explore with Pioneers in Amazon Travel International Expeditions intentionally varies the villages and International Expeditions pioneered travel to the Peruvian tributaries we visit, ensuring you get the most authentic Amazon, and our nature-focused, small-group expedition travel rainforest tour experience remains the standard for Amazon Voyage expedition cruises. IE not only pioneered travel to this region, we still work with many of the naturalist guides that we helped to train. Ensuring a worry-free and educational journey, these expert local naturalist guides — along with an expedition leader — accompany our Amazon Voyage cruises. Each day, these knowledgeable Amazon River travel guides use their decades of learning and guiding experience to serve as lecturers, field guides and friends as you explore the rainforest. -

Suspended Sediment Variability at the Solimões and Negro Confluence Between May 2013 and February 2014
geosciences Communication Suspended Sediment Variability at the Solimões and Negro Confluence between May 2013 and February 2014 Thiago Marinho 1,* ID , Naziano Filizola 2, Jean-Michel Martinez 3, Elisa Armijos 4 and André Nascimento 1 1 Department of Geography, Universidade Federal do Amazonas, 69080-900 Manaus, Brazil; [email protected] 2 Department of Geosciences, Universidade Federal do Amazonas, 69080-900 Manaus, Brazil; nazianofi[email protected] 3 GET, UMR 5563, IRD/CNRS, Université Toulouse 3, 31400 Toulouse, France; [email protected] 4 Instituto Geofísico del Perú—IGP, 15012 Lima, Peru; [email protected] * Correspondence: [email protected]; Tel.: +55-92-993-493-644 Received: 5 July 2018; Accepted: 16 July 2018; Published: 19 July 2018 Abstract: This study focuses on the confluence of two major rivers of the world, the Solimões River (white waters) and Negro River (black waters). Surface suspended sediment samples (SSC) and spectroradiometer taken along transverse profiles at 500 m intervals over a distance of 10 km, as well as satellite images (MODIS) during the hydrological year, were used to follow suspended sediment variability. In January and February, the confluence is dominated by white waters from the Solimões River in the two banks, and in June and July in the right bank by black waters from the Negro River and in the left bank by clear waters from the Solimões River. We found that indirect tools, such as reflectance obtained by spectrometer or MODIS images, can be used to determine surface suspended sediments in a contrasting zone. Keywords: MODIS image; spectrometry; suspended sediments; Negro River; Amazon River; Amazon Basin 1. -

Watershed Technical Analysis
Trout Creek Watershed Stormwater Management Plan SECTION III: WATERSHED TECHNICAL ANALYSIS A. Watershed Modeling An initial step this study of the Trout Creek watershed was the selection of a stormwater simulation model to be used in the analysis. To provide a reasonable estimate of existing flows within the watershed to serve as a baseline for the study of stormwater management and to effectively evaluate the hydrologic response of the watershed to various changes in stormwater management it was necessary to select a model which: • Modeled design storms of various durations and frequencies to produce routed hydrographs which could be combined • Was adaptable to the size of subwatersheds in this study • Could evaluate specific physical characteristics of the rainfall-runoff process • Did not require an excessive amount of input data yet yielded reliable results The model selected for use in this study was the U. S. Army Corps of Engineers, Hydrologic Engineering Center, Hydrologic Modeling System (HEC-HMS) for the following reasons: • It had been developed at the Hydrologic Engineering Center specifically for the analysis of the timing of surface flow contributions to peak rates at various locations in a watershed • Although originally developed as an urban runoff simulation model, data requirements make it easily adaptable to a rural situation • Input parameters provide a flexible calibration process • It has the ability to analyze reservoir or detention basin routing effects • It is accepted by the Pennsylvania Department of Environmental Protection Although other models, such as TR-20, may provide essentially the same results as the HEC-HMS, HMS’s ability to compare subwatershed contributions in a peak flow presentation table make it specifically attractive for this study. -

The Cedar River
Since the 1800’s, Iowa rivers have been designated as CEDARCEDAR either meandered or non- RIVERRIVER meandered. Much of the Cedar River Water Trail is lined with public lands and classified as meandered, meaning that BLACK HAWK COUNTY paddlers may access not only BLACK HAWK the river itself but also the COUNTY river bottom and banks along its course. However, north of Washington Park, the river is classified as non-meandered, meaning that the river bed and surrounding land are owned by the adjacent landowners and paddlers should respect their property rights. BLACK HAWK COUNTY THE CEDAR RIVER - WILDNESS AND CIVILIZATION The Cedar River Water Trail offers a unique paddling experience amongst Iowa’s designated water trails. A journey down the 47 miles of the trail features dramatically contrasting settings, with the wildness of forested bottomlands rich with diverse plant, wildlife, and bird species giving way to historic urban environments. Paddlers are offered an opportunity to explore the downtowns of two major Iowa cities before returning again to the river’s pastoral setting. Traversing the county’s widest stream, its landscape, and lore may feel like time travel at times. From prehistoric times into the present, the Silver maples shade a peaceful backwater of the Cedar River Cedar River valley continues to beckon These trees, typical of bottomland hardwood forest, often grow right up to the water’s to paddlers to explore its natural beauty edge. Quiet back channels are “nurseries” for aquatic wildlife and isolated nesting areas for and cultural treasures. birds and wildlife. Red-spotted purple butterfly Bank swallow nest holes Cedar waxwing gathering nest material CEDAR RIVER WATER TRAIL The Cedar River bisects Black Hawk County diagonally as it continues to erode the landform region called the Iowan Surface. -
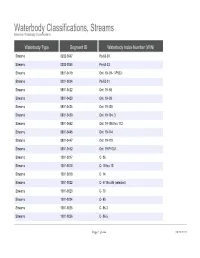
Waterbody Classifications, Streams Based on Waterbody Classifications
Waterbody Classifications, Streams Based on Waterbody Classifications Waterbody Type Segment ID Waterbody Index Number (WIN) Streams 0202-0047 Pa-63-30 Streams 0202-0048 Pa-63-33 Streams 0801-0419 Ont 19- 94- 1-P922- Streams 0201-0034 Pa-53-21 Streams 0801-0422 Ont 19- 98 Streams 0801-0423 Ont 19- 99 Streams 0801-0424 Ont 19-103 Streams 0801-0429 Ont 19-104- 3 Streams 0801-0442 Ont 19-105 thru 112 Streams 0801-0445 Ont 19-114 Streams 0801-0447 Ont 19-119 Streams 0801-0452 Ont 19-P1007- Streams 1001-0017 C- 86 Streams 1001-0018 C- 5 thru 13 Streams 1001-0019 C- 14 Streams 1001-0022 C- 57 thru 95 (selected) Streams 1001-0023 C- 73 Streams 1001-0024 C- 80 Streams 1001-0025 C- 86-3 Streams 1001-0026 C- 86-5 Page 1 of 464 09/28/2021 Waterbody Classifications, Streams Based on Waterbody Classifications Name Description Clear Creek and tribs entire stream and tribs Mud Creek and tribs entire stream and tribs Tribs to Long Lake total length of all tribs to lake Little Valley Creek, Upper, and tribs stream and tribs, above Elkdale Kents Creek and tribs entire stream and tribs Crystal Creek, Upper, and tribs stream and tribs, above Forestport Alder Creek and tribs entire stream and tribs Bear Creek and tribs entire stream and tribs Minor Tribs to Kayuta Lake total length of select tribs to the lake Little Black Creek, Upper, and tribs stream and tribs, above Wheelertown Twin Lakes Stream and tribs entire stream and tribs Tribs to North Lake total length of all tribs to lake Mill Brook and minor tribs entire stream and selected tribs Riley Brook -

Community Report
Community Report Dear Friends, As we refl ect on 2019 so far and look forward to the rest of the Our Mission year, I am thankful for all of those who give their time and talents to make Confl uence Health a place I am proud to be a part of We are dedicated to improving our patients’ every day. Our vision states that Confl uence Health strives “to health by providing safe, high-quality care in become the highest value rural health care system in the nation,” a compassionate and cost-effective manner. About Us and that goal isn’t achieved by accident. Our success is directly impacted by the dedication of our exceptional physicians, nurses, employees, volunteers and leadership teams, who all share a commitment to our patients. Healthy individuals make Our Vision healthy communities, and we understand and embrace the part we play in making that a reality. To become the highest value rural health 4, 000+ employees care system in the nation that improves Additionally, fulfi lling this vision means ensuring the best possible health, quality of life, and is a source of health outcomes at the lowest possible cost, whenever and wherever our patients need care. If we can accomplish that, pride to those who work here. we will uphold our founding principle that a locally controlled, 12 cities directed and integrated health care delivery system best meets the needs of this region. While acknowledging our successes, we understand there is still 12,000 much to do to ensure our community receives the effective and square mile timely care it deserves. -

Holocene Environmental Archaeology of the Yangtze River Valley in China: a Review
land Review Holocene Environmental Archaeology of the Yangtze River Valley in China: A Review Li Wu 1,2,*, Shuguang Lu 1, Cheng Zhu 3, Chunmei Ma 3, Xiaoling Sun 1, Xiaoxue Li 1, Chenchen Li 1 and Qingchun Guo 4 1 Provincial Key Laboratory of Earth Surface Processes and Regional Response in the Yangtze-Huaihe River Basin, School of Geography and Tourism, Anhui Normal University, Wuhu 241002, China; [email protected] (S.L.); [email protected] (X.S.); [email protected] (X.L.); [email protected] (C.L.) 2 State Key Laboratory of Loess and Quaternary Geology, Institute of Earth Environment, Chinese Academy of Sciences, Xi’an 710061, China 3 School of Geograpy and Ocean Science, Nanjing University, Nanjing 210023, China; [email protected] (C.Z.); [email protected] (C.M.) 4 School of Environment and Planning, Liaocheng University, Liaocheng 252000, China; [email protected] * Correspondence: [email protected] Abstract: The Yangtze River Valley is an important economic region and one of the cradles of human civilization. It is also the site of frequent floods, droughts, and other natural disasters. Conducting Holocene environmental archaeology research in this region is of great importance when studying the evolution of the relationship between humans and the environment and the interactive effects humans had on the environment from 10.0 to 3.0 ka BP, for which no written records exist. This Citation: Wu, L.; Lu, S.; Zhu, C.; review provides a comprehensive summary of materials that have been published over the past Ma, C.; Sun, X.; Li, X.; Li, C.; Guo, Q. -

The Effect of Different Confluence Confirmation Strategies on the Obturation of Vertucci Type II Canal: Micro-CT Analysis
Restor Dent Endod. 2021 Feb;46(1):e12 https://doi.org/10.5395/rde.2021.46.e12 pISSN 2234-7658·eISSN 2234-7666 Research Article The effect of different confluence confirmation strategies on the obturation of Vertucci type II canal: micro-CT analysis Seungjae Do , Min-Seock Seo * Department of Conservative Dentistry, Wonkwang University Daejeon Dental Hospital, Daejeon, Korea Received: Apr 8, 2020 Revised: Jun 7, 2020 ABSTRACT Accepted: Jun 17, 2020 Objectives: The present study aims to compare the obturation quality of 2 confluence Do S, Seo MS confirmation techniques in artificial maxillary first premolars showing Vertucci type II root canal configuration. *Correspondence to Min-Seock Seo, DDS, PhD Materials and Methods: Thirty artificial maxillary premolars having Vertucci type II root Associate Professor, Department of canal configuration were made. They were divided into 3 groups according to the confluence Conservative Dentistry, Wonkwang University confirmation technique as follows. Gutta-percha indentation (GPI) group (confluence Daejeon Dental Hospital, 77 Dunsan-ro, Seo- confirmation using a gutta-percha cone and a K file); electronic apex locator (EAL) group gu, Daejeon 35233, Korea. (confluence confirmation using K files and EAL); and no confluence detection (NCD) E-mail: [email protected] group. In the GPI group and the EAL group, shaping and obturation were performed with Copyright © 2021. The Korean Academy of the modified working length (WL). In the NCD group, shaping was performed without WL Conservative Dentistry adjustment and obturation was carried out with an adjusted master cone. Micro-computed This is an Open Access article distributed tomography was used before preparation and after obturation to calculate the percentage under the terms of the Creative Commons of gutta-percha occupied volume (%GPv) and the volume increase in the apical 4 mm. -

Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington
Western Washington University Western CEDAR WWU Graduate School Collection WWU Graduate and Undergraduate Scholarship Winter 1993 Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington Roger G. Bertschi Western Washington University, [email protected] Follow this and additional works at: https://cedar.wwu.edu/wwuet Part of the Geology Commons Recommended Citation Bertschi, Roger G., "Channel Changes and Flood Frequency on the Upper Main Stem of the Nooksack River, Whatcom County, Washington" (1993). WWU Graduate School Collection. 717. https://cedar.wwu.edu/wwuet/717 This Masters Thesis is brought to you for free and open access by the WWU Graduate and Undergraduate Scholarship at Western CEDAR. It has been accepted for inclusion in WWU Graduate School Collection by an authorized administrator of Western CEDAR. For more information, please contact [email protected]. CHANNEL CHANGES AND FLOOD FREQUENCY ON THE UPPER MAIN STEM OF THE NOOKSACK RIVER, WHATCOM COUNTY, WASHINGTON by Roger G. Bertschi Accepted in Partial Completion of the Requirements for the Degree Master of Science Advisory Committee MASTER'S THESIS In presenting this thesis in partial fulfillment of the requirements for a master's degree at Western Washington University, I agree that the Library shall make its copies freely available for inspection. I further agree that extensive copying of this thesis is allowable only for scholarly purposes. It is understood, however, that any copying or publication of this thesis for commercial purposes, or for financial gain, shall not be allowed without mv permission. Signature Date MASTER’S THESIS In presenting this thesis in partial fulfillment of the requirements for a master’s degree at Western Washington University, I grant to Western Washington University the non-exclusive royalty-free right to archive, reproduce, distribute, and display the thesis in any and all forms, including electronic format, via any digital library mechanisms maintained by WWU. -

Modelling Confluence Dynamics in Large Sand-Bed Braided Rivers
Earth Surf. Dynam. Discuss., https://doi.org/10.5194/esurf-2018-85 Manuscript under review for journal Earth Surf. Dynam. Discussion started: 18 December 2018 c Author(s) 2018. CC BY 4.0 License. 1 Modelling confluence dynamics in large sand-bed braided rivers 2 Haiyan Yang1, Zhenhuan Liu2 3 1College of Water Conservancy and Civil Engineering, South China Agricultural 4 University, Guangzhou 510642, China; [email protected] 5 2 Guangdong Provincial Key Laboratory of Urbanization and Geo-simulation, School 6 of Geography and Planning, Sun Yat-sen University, Guangzhou 510275, China 7 Correspondence: [email protected] 8 Abstract 9 Confluences are key morphological nodes in braided rivers where flow converges, 10 creating complex flow patterns and rapid bed deformation. Field survey and laboratory 11 experimental studies have been carried out to investigate the morphodynamic features 12 in individual confluences, but few have investigated the evolution process of 13 confluences in large braided rivers. In the current study a physics-based numerical 14 model was applied to simulate a large lowland braided river dominated by suspended 15 sediment transport, and analyzed the morphologic changes at confluences and their 16 controlling factors. It was found that the confluences in large braided rivers exhibit 17 some dynamic processes and geometric characteristics that are similar to those observed 18 in individual confluences arising from two tributaries. However, they also show some 19 unique characteristics that are result from the influence of the overall braided pattern 20 and especially of neighboring upstream channels. 21 Key words: braided river, numerical model, confluence, dynamics, geometry, scour 22 hole 1 Earth Surf.