In This Lecture, We Will Review the Basic Concepts of Chirality Within Organic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
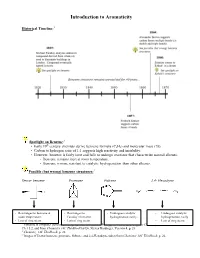
Introduction to Aromaticity
Introduction to Aromaticity Historical Timeline:1 Spotlight on Benzene:2 th • Early 19 century chemists derive benzene formula (C6H6) and molecular mass (78). • Carbon to hydrogen ratio of 1:1 suggests high reactivity and instability. • However, benzene is fairly inert and fails to undergo reactions that characterize normal alkenes. - Benzene remains inert at room temperature. - Benzene is more resistant to catalytic hydrogenation than other alkenes. Possible (but wrong) benzene structures:3 Dewar benzene Prismane Fulvene 2,4- Hexadiyne - Rearranges to benzene at - Rearranges to - Undergoes catalytic - Undergoes catalytic room temperature. Faraday’s benzene. hydrogenation easily. hydrogenation easily - Lots of ring strain. - Lots of ring strain. - Lots of ring strain. 1 Timeline is computer-generated, compiled with information from pg. 594 of Bruice, Organic Chemistry, 4th Edition, Ch. 15.2, and from Chemistry 14C Thinkbook by Dr. Steven Hardinger, Version 4, p. 26 2 Chemistry 14C Thinkbook, p. 26 3 Images of Dewar benzene, prismane, fulvene, and 2,4-Hexadiyne taken from Chemistry 14C Thinkbook, p. 26. Kekulé’s solution: - “snake bites its own tail” (4) Problems with Kekulé’s solution: • If Kekulé’s structure were to have two chloride substituents replacing two hydrogen atoms, there should be a pair of 1,2-dichlorobenzene isomers: one isomer with single bonds separating the Cl atoms, and another with double bonds separating the Cl atoms. • These isomers were never isolated or detected. • Rapid equilibrium proposed, where isomers interconvert so quickly that they cannot be isolated or detected. • Regardless, Kekulé’s structure has C=C’s and normal alkene reactions are still expected. - But the unusual stability of benzene still unexplained. -

II. Stereochemistry 5
B.Sc.(H) Chemistry Semester - II Core Course - III (CC-III) Organic Chemistry - I II. Stereochemistry 5. Physical and Chemical Properties of Stereoisomers Dr. Rajeev Ranjan University Department of Chemistry Dr. Shyama Prasad Mukherjee University, Ranchi 1 Syllabus & Coverage Syllabus II Stereochemistry: Fischer Projection, Newmann and Sawhorse Projection formulae and their interconversions. Geometrical isomerism: cis–trans and syn-anti isomerism, E/Z notations with Cahn Ingold and Prelog (CIP) rules for determining absolute configuration. Optical Isomerism: Optical Activity, Specific Rotation, Chirality/Asymmetry, Enantiomers, Molecules with two or more chiral-centres, Distereoisomers, Meso structures, Racemic mixture. Resolution of Racemic mixtures. Relative and absolute configuration: D/L and R/S designations. Coverage: 1. Types of Isomers : Comparing Structures 2. Optical Activity 3. Racemic Mixtures : Separation of Racemic Mixtures 4. Enantiomeric Excess and Optical Purity 5. Relative and Absolute Configuration 6. Physical and Chemical Properties of Stereoisomers 2 Stereochemistry Types of Isomers Dr. Rajeev Ranjan 3 Stereochemistry Determining the Relationship Between Two Non-Identical Molecules Dr. Rajeev Ranjan 4 Stereochemistry Comparing Structures: Are the structures connected the same? yes no Are they mirror images? Constitutional Isomers yes no Enantiomers Enantiomers Is there a plane of symmetry? All chiral centers will be opposite between them. yes no Meso Diastereomers superimposable Dr. Rajeev Ranjan 5 Stereochemistry Optical Activity: • The chemical and physical properties of two enantiomers are identical except in their interaction with chiral substances. • The physical property that differs is the behavior when subjected to plane-polarized light ( this physical property is often called an optical property). • Plane-polarized (polarized) light is light that has an electric vector that oscillates in a single plane. -

Isomer Distributions of Molecular Weight 247 and 273 Nitro-Pahs in Ambient Samples, NIST Diesel SRM, and from Radical-Initiated Chamber Reactions
Atmospheric Environment 55 (2012) 431e439 Contents lists available at SciVerse ScienceDirect Atmospheric Environment journal homepage: www.elsevier.com/locate/atmosenv Isomer distributions of molecular weight 247 and 273 nitro-PAHs in ambient samples, NIST diesel SRM, and from radical-initiated chamber reactions Kathryn Zimmermann a,1, Roger Atkinson a,1,2,3, Janet Arey a,1,2,*, Yuki Kojima b,4, Koji Inazu b,5 a Air Pollution Research Center, University of California, Riverside, CA 92521, USA b Tokyo Institute of Technology, 4259 Nagatsuta, Midori-ku, Yokohama 226-8502, Japan article info abstract Article history: Molecular weight (mw) 247 nitrofluoranthenes and nitropyrenes and mw 273 nitrotriphenylenes (NTPs), Received 27 December 2011 nitrobenz[a]anthracenes, and nitrochrysenes were quantified in ambient particles collected in Riverside, Received in revised form CA, Tokyo, Japan, and Mexico City, Mexico. 2-Nitrofluoranthene (2-NFL) was the most abundant nitro- 28 February 2012 polycyclic aromatic hydrocarbon (nitro-PAH) in Riverside and Mexico City, and the mw 273 nitro-PAHs Accepted 5 March 2012 were observed in lower concentrations. However, in Tokyo concentrations of 1- þ 2-NTP were more similar to that of 2-NFL. NIST SRM 1975 diesel extract standard reference material was also analyzed to Keywords: examine nitro-PAH isomer distributions, and 12-nitrobenz[a]anthracene was identified for the first time. Nitro-PAH fl Atmospheric reactions The atmospheric formation pathways of nitro-PAHs were studied from chamber reactions of uo- Ambient particles ranthene, pyrene, triphenylene, benz[a]anthracene, and chrysene with OH and NO3 radicals at room Nitrotriphenylenes temperature and atmospheric pressure, with the PAH concentrations being controlled by their vapor pressures. -

Influence of Chirality on the Electromagnetic Wave Electrical and Computer Engineering
Influence of Chirality on the Electromagnetic Wave Propagation: Unbounded Media And Chirowaveguides Priyá Dilipa Gaunço Dessai Dissertation submitted to obtain the Master Degree in Electrical and Computer Engineering Jury President: Professor José Manuel Bioucas Dias Supervisor: Professor Carlos Manuel dos Reis Paiva Co- Supervisor: Professor António Luís da Silva Topa Member Professor Sérgio de Almeida Matos December 2011 Abstract When a chiral medium interacts with the polarization state of an electromag- netic plane wave and couples selectively with either the left or right circularly polarized component, we call this property the optical activity. Since the beginning of the 19th century, the study of complex materials has intensied, and the chiral and bi-isotropic (BI) media have generated one of the most interesting and challenging subjects in the electromagnetic research groups in terms of theoretical problems and potential applications. This dissertation addresses the theoretical interaction between waves and the chiral media. From the study of chiral structures it is possible to observe the eect of the polarization rotation, the propagation modes and the cuto frequencies. The reection and transmission coecients between a simple isotropic media (SIM) and chiral media are also analyzed, as well as the relation between the Brewster angle and the chiral parameter. The BI planar structures are also analyzed for a closed guide, the parallel-plate chirowaveguide, and for a semi-closed guide, the grounded chiroslab. From these structures -

Locally Enhanced and Tunable Optical Chirality in Helical Metamaterials
hv photonics Article Locally Enhanced and Tunable Optical Chirality in Helical Metamaterials Philipp Gutsche 1,*, Raquel Mäusle 1 and Sven Burger 1,2 1 Zuse Institute Berlin, Takustr. 7, 14195 Berlin, Germany; [email protected] (R.M.); [email protected] (S.B.) 2 JCMwave GmbH, Bolivarallee 22, 14050 Berlin, Germany * Correspondence: [email protected]; Tel.: +49-30-841-85-203 Received: 31 October 2016; Accepted: 18 November 2016; Published: 23 November 2016 Abstract: We report on a numerical study of optical chirality. Intertwined gold helices illuminated with plane waves concentrate right and left circularly polarized electromagnetic field energy to sub-wavelength regions. These spots of enhanced chirality can be smoothly shifted in position and magnitude by varying illumination parameters, allowing for the control of light-matter interactions on a nanometer scale. Keywords: metamaterials; optical chirality; computational nano-optics 1. Introduction Helical metamaterials strongly impact the optical response of incident circularly (CPL) and linearly polarized light. They serve as efficient circular polarizers [1] and are candidates for chiral near-field sources [2]. Both, advancement of fabrication techniques [3–5] and design which employs fundamental physical properties [6] have significantly increased the performance of these complex structures. Most experimental, numerical and theoretical studies focus on the far-field response of helical metamaterials. Nevertheless, near-field interaction of light and chiral matter is expected to be enhanced in chiral near-fields. Recently, the quantity of optical chirality has been introduced quantifying this phenomenon [7]. In the weak-coupling regime, the interplay of electromagnetic fields and chiral molecules, which are not superimposable with their mirror image, is directly proportional to this near-field measure [8]. -

Questions & Answers for the New Chemicals Program
Note: Effective January 19, 2016, PMNs must be submitted electronically. Learn more about the new e-PMN requirements. Questions & Answers for the New Chemicals Program (Q&A) U.S. Environmental Protection Agency Office of Pollution Prevention and Toxics Washington, DC 20460 2004 -1- TABLE OF CONTENTS Page 1. GENERAL PROGRAM INFORMATION 100. General ............................................................................................................ 1-1 101. Guidance for Completion of §5 Submission Form ......................................... 1-6 102. Inventory Searches/Bona Fides ....................................................................... 1-17 103. Chemical Identification ................................................................................... 1-22 104. Nomenclature .................................................................................................. 1-26 105. Inventory Issues ................................................................................................ 1-31 106. Review Process ............................................................................................... 1-31 107. Notice of Commencement .............................................................................. 1-33 108. User Fee .......................................................................................................... 1-35 109. Consolidated Notices ...................................................................................... 1-39 110. Joint Submissions .......................................................................................... -

Chirality in Chemical Molecules
Chirality in Chemical Molecules. Molecules which are active in human physiology largely function as keys in locks. The active molecule is then called a ligand and the lock a receptor. The structures of both are highly specific to the degree that if one atom is positioned in a different position than that required by the receptor to be activated, then no stimulation of the receptor or only partial activation can take place. Again in a similar fashion if one tries to open the front door with a key that looks almost the same than the proper key for that lock, one will usually fail to get inside. A simple aspect like a lengthwise groove on the key which is on the left side instead of the right side, can mean that you can not open the lock if your key is the “chirally incorrect” one. The second aspect to understand is which molecules display chirality and which do not. The word chiral comes from the Greek which means “hand-like”. Our hands are mirror images of each other and as such are not identical. If they were, then we would not need a right hand and left hand glove. We can prove that they are not identical by trying to lay one hand on top of the other palms up. When we attempt to do this, we observe that the thumbs and fingers do not lie on top of one another. We say that they are non-super imposable upon one another. Since they are not the same and yet are mirror images of each other, they are said to exhibit chirality. -

Chapter 4: Stereochemistry Introduction to Stereochemistry
Chapter 4: Stereochemistry Introduction To Stereochemistry Consider two of the compounds we produced while finding all the isomers of C7H16: CH3 CH3 2-methylhexane 3-methylhexane Me Me Me C Me H Bu Bu Me Me 2-methylhexane H H mirror Me rotate Bu Me H 2-methylhexame is superimposable with its mirror image Introduction To Stereochemistry Consider two of the compounds we produced while finding all the isomers of C7H16: CH3 CH3 2-methylhexane 3-methylhexane H C Et Et Me Pr Pr 3-methylhexane Me Me H H mirror Et rotate H Me Pr 2-methylhexame is superimposable with its mirror image Introduction To Stereochemistry Consider two of the compounds we produced while finding all the isomers of C7H16: CH3 CH3 2-methylhexane 3-methylhexane .Compounds that are not superimposable with their mirror image are called chiral (in Greek, chiral means "handed") 3-methylhexane is a chiral molecule. .Compounds that are superimposable with their mirror image are called achiral. 2-methylhexane is an achiral molecule. .An atom (usually carbon) with 4 different substituents is called a stereogenic center or stereocenter. Enantiomers Et Et Pr Pr Me CH3 Me H H 3-methylhexane mirror enantiomers Et Et Pr Pr Me Me Me H H Me H H Two compounds that are non-superimposable mirror images (the two "hands") are called enantiomers. Introduction To Stereochemistry Structural (constitutional) Isomers - Compounds of the same molecular formula with different connectivity (structure, constitution) 2-methylpentane 3-methylpentane Conformational Isomers - Compounds of the same structure that differ in rotation around one or more single bonds Me Me H H H Me H H H H Me H Configurational Isomers or Stereoisomers - Compounds of the same structure that differ in one or more aspects of stereochemistry (how groups are oriented in space - enantiomers or diastereomers) We need a a way to describe the stereochemistry! Me H H Me 3-methylhexane 3-methylhexane The CIP System Revisited 1. -

Electrophilic Mercuration and Thallation of Benzene and Substituted Benzenes in Trifluoroacetic Acid Solution* (Electrophilic Substitution/Selectivity) GEORGE A
Proc. Natl. Acad. Sci. USA Vol. 74, No. 10, pp. 4121-4125, October 1977 Chemistry Electrophilic mercuration and thallation of benzene and substituted benzenes in trifluoroacetic acid solution* (electrophilic substitution/selectivity) GEORGE A. OLAH, IWAO HASHIMOTOt, AND HENRY C. LINtt Institute of Hydrocarbon Chemistry, Department of Chemistry, University of Southern California, Los Angeles, California 90007; and the Department of Chemistry, Case Western Reserve University, Cleveland, Ohio 44101 Contributed by George A. Olah, July 18, 1977 ABSTRACT The mercuration and thallation of benzene and iments at 250 and quenched the mixtures after 5 min, whereas substituted benzenes was studied with mercuric and thallic Brown and Nelson carried out the reaction for 6.5 hr. In sub- trifluoroacetate, respectively, in trifluoroacetic acid. With the in view of the importance of both the mechanistic shortest reaction time (1 sec) at 00, the relative rate of mercu- sequent work, ration of toluene compared to that of benzene was 17.5, with and practical implications of the orientation-rate correlation, the isomer distribution in toluene of: ortho, 17.4%; meta, 5.9%; Brown and McGary (7), carried out a more detailed study of and para, 76.7%. The isomer distribution in toluene varied with the mercuration reaction. They concluded: "A redetermination the reaction time, significantly more at 25° than at 00. The of the isomer distributions and relative rates indicates excellent competitive thallation of benzene and toluene with thallic tri- agreement with the linear relationship of orientation and rel- fluoroacetate in trifluoroacetic acid at 150 showed the relative ative however, that the isomer distri- rate, toluene/benzene, to be 33, with the isomer distribution in rate." They recognized, toluene of: ortho, 9.5%; meta, 5.5%; and para, 85.0%. -

Cis-Trans Isomerism 2
Isomerism Isomerism • These compounds possess the same molecular formula but differ from each other in physical or chemical properties, and are called Isomers and the phenomenon is termed Isomerism • Since isomers have the same molecular formula, the difference in their properties must be due to different modes of combination or arrangement of atoms within the molecule. There are two main types of isomerism • (i) Structural Isomerism and (ii) Stereoisomerism. Structural Isomerism • When the isomerism is simply due to difference in the arrangement of atoms within the molecule without any reference to space, the phenomenon is termed Structural Isomerism. In other words, the structural isomers are compounds that have the same molecular formula but different structural formulas. Types of Structural Isomerism 1. Chain Isomerism 2. Position Isomerism 3. Functional Isomerism 4. Metamersim 5. Tautomerism Stereoisomerism • When isomerism is caused by the different arrangements of atoms or groups in space, the phenomenon is called Stereoisomerism (Greek, Stereos = occupying space). • The stereoisomers have the same structural formulas but differ in arrangement of atoms in space. In other words, stereoisomerism is exhibited by such compounds which have identical molecular structure but different configurations. Configuration refers to the 3-dimentional arrangement of atoms that characterizes a particular compound. Types of Stereoisomerism Stereoisomerism is of two types 1. Geometrical or Cis-Trans Isomerism 2. Optical Isomerism Structural Isomerism Chain Isomerism This type of isomerism arises from she difference in the structure of carbon chain which forms the nucleus of the molecule. It is, therefore, named as Chain or Nuclear Isomerism. Chain isomers have the same molecular formula but differ in the order in which the carbon atoms are bonded to each other For example, there are known two butanes which have the same molecular formula (C4H10) but differ in the structure of the carbon chains in their molecules. -

The Power of Crowding for the Origins of Life
Orig Life Evol Biosph (2014) 44:307–311 DOI 10.1007/s11084-014-9382-5 ORIGIN OF LIFE The Power of Crowding for the Origins of Life Helen Greenwood Hansma Received: 2 October 2014 /Accepted: 2 October 2014 / Published online: 14 January 2015 # Springer Science+Business Media Dordrecht 2015 Abstract Molecular crowding increases the likelihood that life as we know it would emerge. In confined spaces, diffusion distances are shorter, and chemical reactions produce fewer and more regular products. Crowding will occur in the spaces between Muscovite mica sheets, which has many advantages as a site for life’s origins. Keywords Muscovite mica . Molecular crowding . Origin of life . Mechanochemistry. Abiogenesis . Chemical confinement effects . Chirality. Protocells Cells are crowded. Protein molecules in cells are typically so close to each other that there is room for only one protein molecule between them (Phillips, Kondev et al. 2008). This is nothing like a dilute ‘prebiotic soup.’ Therefore, by analogy with living cells, the origins of life were probably also crowded. Molecular Confinement Effects Many chemical reactions are limited by the time needed for reactants to diffuse to each other. Shorter distances speed up these reactions. Molecular complementarity is another principle of life in which pairs or groups of molecules form specific interactions (Root-Bernstein 2012). Current examples are: enzymes & substrates & cofactors; nucleic acid base pairs; antigens & antibodies; nucleic acid - protein interactions. Molecular complementarity is likely to have been involved at life’s origins and also benefits from crowding. Mineral surfaces are a likely place for life’s origins and for formation of polymeric molecules (Orgel 1998). -

Alkenes and Alkynes
02/21/2019 CHAPTER FOUR Alkenes and Alkynes H N O I Cl C O C O Cl F3C C Cl C Cl Efavirenz Haloprogin (antiviral, AIDS therapeutic) (antifungal, antiseptic) Chapter 4 Table of Content * Unsaturated Hydrocarbons * Introduction and hybridization * Alkenes and Alkynes * Benzene and Phenyl groups * Structure of Alkenes, cis‐trans Isomerism * Nomenclature of Alkenes and Alkynes * Configuration cis/trans, and cis/trans Isomerism * Configuration E/Z * Physical Properties of Hydrocarbons * Acid‐Base Reactions of Hydrocarbons * pka and Hybridizations 1 02/21/2019 Unsaturated Hydrocarbons • Unsaturated Hydrocarbon: A hydrocarbon that contains one or more carbon‐carbon double or triple bonds or benzene‐like rings. – Alkene: contains a carbon‐carbon double bond and has the general formula CnH2n. – Alkyne: contains a carbon‐carbon triple bond and has the general formula CnH2n‐2. Introduction Alkenes ● Hydrocarbons containing C=C ● Old name: olefins • Steroids • Hormones • Biochemical regulators 2 02/21/2019 • Alkynes – Hydrocarbons containing C≡C – Common name: acetylenes Unsaturated Hydrocarbons • Arene: benzene and its derivatives (Ch 9) 3 02/21/2019 Benzene and Phenyl Groups • We do not study benzene and its derivatives until Chapter 9. – However, we show structural formulas of compounds containing a phenyl group before that time. – The phenyl group is not reactive under any of the conditions we describe in chapters 5‐8. Structure of Alkenes • The two carbon atoms of a double bond and the four atoms bonded to them lie in a plane, with bond angles of approximately 120°. 4 02/21/2019 Structure of Alkenes • Figure 4.1 According to the orbital overlap model, a double bond consists of one bond formed by overlap of sp2 hybrid orbitals and one bond formed by overlap of parallel 2p orbitals.