Microbial Communities in Coastal Sediments: Structure and Functions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ecosystem Structure and Function. Dr
TOPIC: - ECOSYSTEM STRUCTURE AND FUNCTION. DR. ABHAY KRISHNA SINGH PAPER NAME: - ENVIRONMENTAL GEOGRAPHY SUBJECT: - GEOGRAPHY SEMESTER: - M.A. –IV PAPER CODE: - (GEOG. 403) UNIVERSITY DEPARTMENT OF GEOGRAPHY, DR. SHYMA PRASAD MUKHERJEE UNIVERSITY, RANCHI. Environmental Sciences INTRODUCTION: - All organisms need energy to perform the essential functions such as maintenance, growth, repair, movement, locomotion and reproduction; all of these processes require energy expenditure. The ultimate source of energy for all ecological systems is Sun. The solar energy is captured by the green plants (primary producers or autotrophs) and transformed into chemical energy and bound in glucose as potential energy during the process of photosynthesis. In this stored form, other organisms take the energy and pass it on further to other organisms. During this process, a reasonable proportion of energy is lost out of the living system. The whole process is called flow of energy in the ecosystem. It is the amount of energy that is received and transferred from organism to organism in an ecosystem that modulates the ecosystem structure. Without autotrophs, there would be no energy available to all other organisms that lack the capability of fixing light energy. A fraction i.e. about 1/50 millionth of the total solar radiation reaches the earth’s atmosphere. About 34% of the sunlight reaching the earth’s atmosphere is reflected back into the atmosphere, 10% is held by ozone layer, water vapors and other atmospheric gases. The remaining 56% sunlight reaches the earth’s surface. Only a fraction of this energy reaching the earth’s surface (1 to 5%) is used by green plants for photosynthesis and the rest is absorbed as heat by ground vegetation or water. -

7.014 Handout PRODUCTIVITY: the “METABOLISM” of ECOSYSTEMS
7.014 Handout PRODUCTIVITY: THE “METABOLISM” OF ECOSYSTEMS Ecologists use the term “productivity” to refer to the process through which an assemblage of organisms (e.g. a trophic level or ecosystem assimilates carbon. Primary producers (autotrophs) do this through photosynthesis; Secondary producers (heterotrophs) do it through the assimilation of the organic carbon in their food. Remember that all organic carbon in the food web is ultimately derived from primary production. DEFINITIONS Primary Productivity: Rate of conversion of CO2 to organic carbon (photosynthesis) per unit surface area of the earth, expressed either in terns of weight of carbon, or the equivalent calories e.g., g C m-2 year-1 Kcal m-2 year-1 Primary Production: Same as primary productivity, but usually expressed for a whole ecosystem e.g., tons year-1 for a lake, cornfield, forest, etc. NET vs. GROSS: For plants: Some of the organic carbon generated in plants through photosynthesis (using solar energy) is oxidized back to CO2 (releasing energy) through the respiration of the plants – RA. Gross Primary Production: (GPP) = Total amount of CO2 reduced to organic carbon by the plants per unit time Autotrophic Respiration: (RA) = Total amount of organic carbon that is respired (oxidized to CO2) by plants per unit time Net Primary Production (NPP) = GPP – RA The amount of organic carbon produced by plants that is not consumed by their own respiration. It is the increase in the plant biomass in the absence of herbivores. For an entire ecosystem: Some of the NPP of the plants is consumed (and respired) by herbivores and decomposers and oxidized back to CO2 (RH). -

Model-Based Analysis of the Energy Fluxes and Trophic Structure of a Portunus Trituberculatus Polyculture Ecosystem
Vol. 9: 479–490, 2017 AQUACULTURE ENVIRONMENT INTERACTIONS Published December 5 https://doi.org/10.3354/aei00247 Aquacult Environ Interact OPENPEN ACCESSCCESS Model-based analysis of the energy fluxes and trophic structure of a Portunus trituberculatus polyculture ecosystem Jie Feng1, Xiang-Li Tian1,*, Shuang-Lin Dong1, Rui-Peng He1, Kai Zhang1, Dong-Xu Zhang1, Qing-Qi Zhang2 1The Key Laboratory of Mariculture, Ministry of Education, Fisheries College, Ocean University of China, Qingdao 266003, PR China 2Marine Fishery Technology Guiding Office of Ganyu, Lianyungang 222100, PR China ABSTRACT: We constructed a quantitative Ecopath model of a trophic network to evaluate the energy flow and properties in a polyculture ecosystem containing 4 species (swimming crab Por- tunus trituberculatus, white shrimp Litopenaeus vannamei, short-necked clam Ruditapes philip- pinarum, and redlip mullet Liza haematochila) over a 90 d experimental period. The model con- tained 10 consumers, 4 detritus groups, and 4 primary producers. Ecotrophic efficiency values indicated that the system had high energy utilization efficiency. However, benthic bacteria con- verted the largest amount of energy back to the detritus groups, which had the lowest ecotrophic efficiency (0.01). When aggregating the network to discrete trophic levels (TLs), most of the throughput and biomass of the system were distributed on the first 2 TLs; consequently, there was high energy transfer efficiency between TL I and II (81.98%). The trophic flow of this ecosystem was dominated by energy that originated from the detritus groups (73.77%). Imported artificial food was particularly important for the trophic flow of the total ecosystem, contributing 31.02% to total system consumption. -
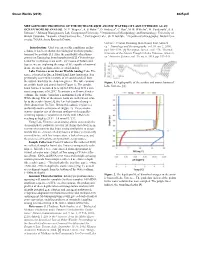
Metagenomic Profiling of the Methane-Rich Anoxic Waters of Lake Untersee As an Ocean Worlds Analog
Ocean Worlds (2019) 6025.pdf METAGENOMIC PROFILING OF THE METHANE-RICH ANOXIC WATERS OF LAKE UNTERSEE AS AN OCEAN WORLDS ANALOG. N. Y. Wagner1, A. S. Hahn2,3, D. Andersen4, C. Roy5, M. B. Wilhelm6, M. Vanderwilt1, S. S. Johnson1, 1 Johnson Biosignatures Lab, Georgetown University, 2 Department of Microbiology and Immunology, University of British Columbia, 3 Koonkie Cloud Services Inc., 4 Carl Sagan Center, SETI Institute, 5 Department of Geography, McGill Uni- versity, 6NASA Ames Research Center. Untersee, Central Dronning Maud Land, East Antarcti- Introduction: Under ocean worlds conditions on En- ca.” Limnology and Oceanography, vol. 51, no. 2, 2006, celadus, it has been shown that biological methane produc- pp.1180–1194, [4] Bevington, James, et al. “The Thermal tion may be possible [1]. Also, the possibility of methano- Structure of the Anoxic Trough in Lake Untersee, Antarcti- genesis on Europa has been hypothesized [2]. Given the po- ca.”Antarctic Science, vol. 30, no. 6, 2018, pp. 333–344. tential for methanogenesis on the icy moons of Saturn and Jupiter, we are exploring the range of life capable of survival in an extremely methane-rich terrestrial analog. Lake Untersee as an Ocean Worlds Analog: Lake Un- tersee is located in Queen Maud Land, East Antarctica. It is perennially covered in 3 meters of ice and closed off from the outside world by the Anuchin glacier. The lake contains Figure 1. Depth profile of the aerobic and anoxic basins of an aerobic basin and anoxic basin (Figure 1). The aerobic Lake Untersee [4]. basin has been measured to be up to 169m deep with a con- stant temperature of 0.25˚C. -

Algal Toxic Compounds and Their Aeroterrestrial, Airborne and Other Extremophilic Producers with Attention to Soil and Plant Contamination: a Review
toxins Review Algal Toxic Compounds and Their Aeroterrestrial, Airborne and other Extremophilic Producers with Attention to Soil and Plant Contamination: A Review Georg G¨аrtner 1, Maya Stoyneva-G¨аrtner 2 and Blagoy Uzunov 2,* 1 Institut für Botanik der Universität Innsbruck, Sternwartestrasse 15, 6020 Innsbruck, Austria; [email protected] 2 Department of Botany, Faculty of Biology, Sofia University “St. Kliment Ohridski”, 8 blvd. Dragan Tsankov, 1164 Sofia, Bulgaria; mstoyneva@uni-sofia.bg * Correspondence: buzunov@uni-sofia.bg Abstract: The review summarizes the available knowledge on toxins and their producers from rather disparate algal assemblages of aeroterrestrial, airborne and other versatile extreme environments (hot springs, deserts, ice, snow, caves, etc.) and on phycotoxins as contaminants of emergent concern in soil and plants. There is a growing body of evidence that algal toxins and their producers occur in all general types of extreme habitats, and cyanobacteria/cyanoprokaryotes dominate in most of them. Altogether, 55 toxigenic algal genera (47 cyanoprokaryotes) were enlisted, and our analysis showed that besides the “standard” toxins, routinely known from different waterbodies (microcystins, nodularins, anatoxins, saxitoxins, cylindrospermopsins, BMAA, etc.), they can produce some specific toxic compounds. Whether the toxic biomolecules are related with the harsh conditions on which algae have to thrive and what is their functional role may be answered by future studies. Therefore, we outline the gaps in knowledge and provide ideas for further research, considering, from one side, Citation: G¨аrtner, G.; the health risk from phycotoxins on the background of the global warming and eutrophication and, ¨а Stoyneva-G rtner, M.; Uzunov, B. -
![3.2 Energy Flows Through Ecosystems [Notes/Highlighting]](https://docslib.b-cdn.net/cover/9150/3-2-energy-flows-through-ecosystems-notes-highlighting-899150.webp)
3.2 Energy Flows Through Ecosystems [Notes/Highlighting]
Printed Page 60 3.2 Energy flows through ecosystems [Notes/Highlighting] To understand how ecosystems function and how to best protect and manage them, ecosystem ecologists study not only the biotic and abiotic components that define an ecosystem, but also the processes that move energy and matter within it. Plants absorb energy directly from the Sun. That energy is then spread throughout an ecosystem as herbivores (animals that eat plants) feed on plants and carnivores (animals that eat other animals) feed on herbivores. Consider the Serengeti Plain in East Africa, shown in FIGURE 3.3. There are millions of herbivores, such as zebras and wildebeests, in the Serengeti ecosystem, but far fewer carnivores, such as lions (Panthera leo) and cheetahs (Acinonyx jubatus), that feed on those herbivores. In accordance with the second law of thermodynamics, when one organism consumes another, not all of the energy in the consumed organism is transferred to the consumer. Some of that energy is lost as heat. Therefore, all the carnivores in an area contain less energy than all the herbivores in the same area because all the energy going to the carnivores must come from the animals they eat. To better understand these energy relationships, let’s trace this energy flow in more detail. Figure 3.3 Serengeti Plain of Africa. The Serengeti ecosystem has more plants than herbivores, and more herbivores than carnivores. Previous Section | Next Section 3.2.1 Photosynthesis and Respiration Printed Page 60 [Notes/Highlighting] Nearly all of the energy that powers ecosystems comes from the Sun as solar energy, which is a form of kinetic energy. -

AP Biology Flash Review Is Designed to Help Howyou Prepare to Use Forthis and Book Succeed on the AP Biology Exam
* . .AP . BIOLOGY. Flash review APBIOL_00_ffirs_i-iv.indd 1 12/20/12 9:54 AM OTHER TITLES OF INTEREST FROM LEARNINGEXPRESS AP* U.S. History Flash Review ACT * Flash Review APBIOL_00_ffirs_i-iv.indd 2 12/20/12 9:54 AM AP* BIOLOGY . Flash review ® N EW YORK APBIOL_00_ffirs_i-iv.indd 3 12/20/12 9:54 AM The content in this book has been reviewed and updated by the LearningExpress Team in 2016. Copyright © 2012 LearningExpress, LLC. All rights reserved under International and Pan American Copyright Conventions. Published in the United States by LearningExpress, LLC, New York. Printed in the United States of America 987654321 First Edition ISBN 978-1-57685-921-6 For more information or to place an order, contact LearningExpress at: 2 Rector Street 26th Floor New York, NY 10006 Or visit us at: www.learningexpressllc.com *AP is a registered trademark of the College Board, which was not involved in the production of, and does not endorse, this product. APBIOL_00_ffirs_i-iv.indd 4 12/20/12 9:54 AM Contents 1 . .. 11 IntRoDUCtIon 57 . ... A. 73 . ... B. 131 . ... C. 151 . .... D. 175 . .... e. 183 . .... F. 205 . .... G. 225 . .... H. 245 . .... I. 251 . .... K. 267 . .... L. 305 . .... M. [ v ] . .... n. APBIOL_00_fcont_v-viii.indd 5 12/20/12 9:55 AM 329 343 . .... o. 411 . .... P. 413 . .... Q. 437 . .... R. 489 . .... s. 533 . .... t. 533 . .... U. 539 . .... V. 541 . .... X. .... Z. [ vi ] APBIOL_00_fcont_v-viii.indd 6 12/20/12 9:55 AM * . .AP . BIOLOGY. FLAsH.ReVIew APBIOL_00_fcont_v-viii.indd 7 12/20/12 9:55 AM Blank Page 8 APBIOL_00_fcont_v-viii.indd 8 12/20/12 9:55 AM IntroductIon The AP Biology exam tests students’ knowledge Aboutof core themes, the AP topics, Biology and concepts Exam covered in a typical high school AP Biology course, which offers students the opportunity to engage in college-level biology study. -

Chapter 15 Communities and Ecosystems Rosech15 0104043 437-474 2P 11/18/04 3:07 PM Page 439
RoseCh15_0104043_437-474_2p 11/18/04 2:32 PM Page 437 15 The feeding relationships between species can often be complicated. Communities and Ecosystems hen scientists first began studying bio- dioxide levels, which are covered in Chapter 16 logical communities, they were so fasci- (The Biosphere and the Physical Environment). Wnated with the interactions and The coordination and integration of biological dependencies between species that they saw the bi- communities has vast implications for the Earth. ological community as a superorganism. Whole For this reason, there are few biological topics as species were viewed as organs that performed spe- important for the future of life on Earth as the func- cific functions for the complete ecological superor- tioning of ecosystems. In this chapter, we survey ganism. The integration and communication how ecosystems function, from the flow of energy in between these “organs” was thought to be deliber- Module 15.1 (Energy Flow) and the recycling of nu- ate and well tuned. One way to think of this idea is trients in Module 15.15 (Ecosystems) to the porten- to imagine a stitched-together Frankenstein, each tous problem of the fragility of ecosystems. In sewn-on body part a distinct species. Modules 15.8 (Community Organization) and 15.4 Today biologists find the analogy between bio- (Equilibrium and Nonequilibrium Communities), logical communities and organisms superficial. To we consider the factors that determine the number be sure, there are populations within communities of species in a community. Surprisingly, in some that are highly dependent on each other. And it is communities predation and environmental distur- also true that biological communities and their bance may promote increased species diversity. -

Technical Efficiency, Ecological Efficiency and Grassland Ecological
Technical Efficiency, Ecological Efficiency and Grassland Ecological Performance of Grazing in China Wei Huang1*, Bernhard Brümmer1, Lynn Huntsinger2 1 Department for agricultural economics and rural development, University of Goettingen, Germany 2 Department of environmental science, policy, and management, University of California at Berkeley, US * Correspondent email: [email protected] Paper prepared for presentation at the EAAE 2014 Congress ‘Agri-Food and Rural Innovations for Healthier Societies’ August 26 to 29, 2014 Ljubljana, Slovenia Copyright 2014 by authors. All rights reserved. Readers may make verbatim copies of this document for non-commercial purposes by any means, provided that this copyright notice appears on all such copies Abstract: Incorporating the ecological variable of grassland Net Primary Productivity (NPP) into the production function - to be representative of grassland quality - is a new step toward the ecological efficiency analysis under the field of productivity and efficiency analysis. We measure the technical efficiency, ecological performance indicator and ecological efficiency of grazing using a multi-outputs and multi-inputs stochastic input-oriented distance function. The average technical efficiency is estimated to be 0.90 when taking grassland NPP into account, implying that cost of grazing inputs can be decreased by 10% without any deduction of outputs. The ecological efficiency is estimated to be 0.83 and the average ecological performance indicator is 0.17. Key words: Technical efficiency; Ecological efficiency; Ecological performance indicator; Net primary productivity (NPP); input distance function. 1. Introduction The concerns about environmental problems pushed by local economic development in developing countries received a lot of attention in recent years; as one of the main land use types on earth is grassland, the relationship between environmental problems caught by inappropriate grassland use and local economic development became a popular topic. -

Secondary Production and Ecological Efficiencies A.R.E
SECONDARY PRODUCTION AND ECOLOGICAL EFFICIENCIES A.R.E. Sinclair - Biology 302 1. Pn goes either to decomposers or herbivores. The latter consume (C) some. Of this part is passed out of the animal as urinary (U) or faecal (F) energy, and the rest is absorbed through the gut as assimilated (A) energy. This is used for body maintenance (=respiration R) and production (P). 2. A can be measured by a) lab studies of C,F,U since A = C - F - U, b) lab studies of R and field measures of P since A = R+P 3. R is affected by Basal Metabolic Rate (kcal/day). In mammals this BMR = 70*(Body Wt in kg)0.75 . Found by Brody (1945), Kleiber (1947). 4. Although absolute energy requirements go up with body size the energy per unit weight goes down with increasing body size because the surface area / volume ratio declines. Thus, heat loss/kg body weight also declines. This applies to both homeotherms (warm blooded animals) and poikilotherms (cold blooded). Efficiency within a trophic level 5. Production Efficiency is the ratio of P to A. In poikilotherms more A goes to P in short lived species (<2yrs) because less is needed for R in nonreproductive periods, i.e. they have higher growth efficiency. 6. Respiration loss is the ratio of R to A. In carnivores more A goes to R due to higher searching rates and other activities. Thus, P/A declines with higher trophic levels. 7. More A goes to R in homeotherms than poikilotherms. 8. Assimilation Efficiency (=Digestibility) is the ratio of A to I. -

Productivity Significant Ideas
2.3 Flows of Energy & Matter - Productivity Significant Ideas • Ecosystems are linked together by energy and matter flow • The Sun’s energy drives these flows and humans are impacting the flows of energy and matter both locally and globally Knowledge & Understandings • As solar radiation (insolation) enters the Earth’s atmosphere some energy becomes unavailable for ecosystems as the energy absorbed by inorganic matter or reflected back into the atmosphere. • Pathways of radiation through the atmosphere involve the loss of radiation through reflection and absorption • Pathways of energy through an ecosystem include: • Conversion of light to chemical energy • Transfer of chemical energy from one trophic level to another with varying efficiencies • Overall conversion of UB and visible light to heat energy by the ecosystem • Re-radiation of heat energy to the atmosphere. Knowledge & Understandings • The conversion of energy into biomass for a given period of time is measured by productivity • Net primary productivity (NPP) is calculated by subtracting respiratory losses (R) from gross primary productivity (GPP) NPP = GPP – R • Gross secondary productivity (GSP) is the total energy/biomass assimulated by consumers and is calculated by subtracting the mass of fecal loss from the mass of food eaten. GSP = food eaten – fecal loss • Net secondary productivity (NSP) is calculated by subtracting the respiratory losses (R) from GSP. NSP=GSP - R Applications and Skills • Analyze quantitative models of flows of energy and matter • Construct quantitative -

Extremophile Hunt Begins 8 February 2008
Extremophile Hunt Begins 8 February 2008 Hoover, "is that you don't have to have a 'Goldilocks' zone with perfect temperature, a certain pH level, and so forth, for life to thrive." Researchers have found microbes living in ice, in boiling water, in nuclear reactors. These "strange" extremophiles may in fact be the norm for life elsewhere in the cosmos. "With our research this year, we hope to identify some new limits for life in terms of temperature and pH levels. This will help us decide where to search for life on other planets and how to recognize alien life if we actually find it." Spirochaeta americana, extreme-loving microbes from California's Mono Lake. Hoover et al discovered them Hoover has already made some new friends in cold during a previous extremophile-hunting expedition. places. Earlier Hoover teams have found new species and genera of anaerobic microbial extremophiles in the ice and permafrost of Alaska, Siberia, Patagonia, and Antarctica. A team of scientists has just left the country to explore a very strange lake in Antarctica; it is filled "I found one extremophile in penguin guano," with, essentially, extra-strength laundry detergent. recalls Hoover. "When I stooped to pick it up, Jim No, the researchers haven't spilled coffee on their Lovell, my research partner then, said, 'What the lab coats. They are hunting for extremophiles -- heck are you doing now, Richard?' But it paid off." tough little creatures that thrive in conditions too extreme for most other living things. Most incredible, though, was the revelation a few years ago that some extremophiles the researchers Antarctica's Lake Untersee, fed by glaciers, always found in an Alaskan tunnel actually came to life as covered with ice, and very alkaline, is one of the the ice around them melted.