Jurassic Cretaceous
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Middle Jurassic of Western and Northern Europe: Its Subdivisions, Geochronology and Correlations
The Middle Jurassic of western and northern Europe: its subdivisions, geochronology and correlations John H. Callomon The palaeogeographic settings of Denmark and East Greenland during the Middle Jurassic are outlined. They lay in the widespread epicontinental seas that covered much of Europe in the post-Triassic transgression. It was a period of continuing eustatic sea-level rise, with only distant connections to world oceans: to the Pacific, via the narrow Viking Straits between Greenland and Norway and hence the arctic Boreal Sea to the north; and to the subtropical Tethys, via some 1200 km of shelf-seas to the south. The sedimentary history of the region was strongly influenced by two factors: tectonism and climate. Two modes of tectonic movement governed basinal evolution: crustal extension lead- ing to subsidence through rifting, such as in the Viking and Central Grabens of the North Sea; and subcrustal thermal upwelling, leading to domal uplift and the partition of marine basins through emergent physical barriers, as exemplified by the Central North Sea Dome with its associated volcanics. The climatic gradient across the 30º of temperate latitude spanned by the European seas governed biotic diversity and biogeography, finding expression in rock-forming biogenic carbonates that dominate sediments in the south and give way to largely siliciclastic sediments in the north. Geochronology of unrivalled finesse is provided by standard chronostratigraphy based on the biostratigraphy of ammonites. The Middle Jurassic saw the onset of considerable bioprovincial endemisms in these guide-fossils, making it necessary to construct parallel standard zonations for Boreal, Subboreal or NW European and Submediterranean Provinces, of which the NW European zonation provides the primary international standard. -

Cretaceous Boundary in Western Cuba (Sierra De Los Órganos)
GEOLOGICA CARPATHICA, JUNE 2013, 64, 3, 195—208 doi: 10.2478/geoca-2013-0014 Calpionellid distribution and microfacies across the Jurassic/ Cretaceous boundary in western Cuba (Sierra de los Órganos) RAFAEL LÓPEZ-MARTÍNEZ1, , RICARDO BARRAGÁN1, DANIELA REHÁKOVÁ2 and JORGE LUIS COBIELLA-REGUERA3 1Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, C.P. 04510, México D.F., México; [email protected] 2Comenius University, Faculty of Natural Sciences, Department of Geology and Paleontology, Mlynská dolina G, 842 15 Bratislava, Slovak Republic; [email protected] 3Departamento de Geología, Universidad de Pinar del Río, Martí # 270, Pinar del Río, C.P. 20100, Cuba (Manuscript received May 21, 2012; accepted in revised form December 11, 2012) Abstract: A detailed bed-by-bed sampled stratigraphic section of the Guasasa Formation in the Rancho San Vicente area of the “Sierra de los Órganos”, western Cuba, provides well-supported evidence about facies and calpionellid distribution across the Jurassic/Cretaceous boundary. These new data allowed the definition of an updated and sound calpionellid biozonation scheme for the section. In this scheme, the drowning event of a carbonate platform displayed by the facies of the San Vicente Member, the lowermost unit of the section, is dated as Late Tithonian, Boneti Subzone. The Jurassic/Cretaceous boundary was recognized within the facies of the overlying El Americano Member on the basis of the acme of Calpionella alpina Lorenz. The boundary is placed nearly six meters above the contact between the San Vicente and the El Americano Members, in a facies linked to a sea-level drop. The recorded calpionellid bioevents should allow correlations of the Cuban biozonation scheme herein proposed, with other previously published schemes from distant areas of the Tethyan Domain. -

Asteroid Impact, Not Volcanism, Caused the End-Cretaceous Dinosaur Extinction
Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction Alfio Alessandro Chiarenzaa,b,1,2, Alexander Farnsworthc,1, Philip D. Mannionb, Daniel J. Luntc, Paul J. Valdesc, Joanna V. Morgana, and Peter A. Allisona aDepartment of Earth Science and Engineering, Imperial College London, South Kensington, SW7 2AZ London, United Kingdom; bDepartment of Earth Sciences, University College London, WC1E 6BT London, United Kingdom; and cSchool of Geographical Sciences, University of Bristol, BS8 1TH Bristol, United Kingdom Edited by Nils Chr. Stenseth, University of Oslo, Oslo, Norway, and approved May 21, 2020 (received for review April 1, 2020) The Cretaceous/Paleogene mass extinction, 66 Ma, included the (17). However, the timing and size of each eruptive event are demise of non-avian dinosaurs. Intense debate has focused on the highly contentious in relation to the mass extinction event (8–10). relative roles of Deccan volcanism and the Chicxulub asteroid im- An asteroid, ∼10 km in diameter, impacted at Chicxulub, in pact as kill mechanisms for this event. Here, we combine fossil- the present-day Gulf of Mexico, 66 Ma (4, 18, 19), leaving a crater occurrence data with paleoclimate and habitat suitability models ∼180 to 200 km in diameter (Fig. 1A). This impactor struck car- to evaluate dinosaur habitability in the wake of various asteroid bonate and sulfate-rich sediments, leading to the ejection and impact and Deccan volcanism scenarios. Asteroid impact models global dispersal of large quantities of dust, ash, sulfur, and other generate a prolonged cold winter that suppresses potential global aerosols into the atmosphere (4, 18–20). These atmospheric dinosaur habitats. -

Non-Invasive Imaging Methods Applied to Neo- and Paleo-Ontological Cephalopod Research
Biogeosciences, 11, 2721–2739, 2014 www.biogeosciences.net/11/2721/2014/ doi:10.5194/bg-11-2721-2014 © Author(s) 2014. CC Attribution 3.0 License. Non-invasive imaging methods applied to neo- and paleo-ontological cephalopod research R. Hoffmann1, J. A. Schultz2, R. Schellhorn2, E. Rybacki3, H. Keupp4, S. R. Gerden1, R. Lemanis1, and S. Zachow5 1Institut für Geologie, Mineralogie und Geophysik, Ruhr Universität Bochum, Universitätsstrasse 150, 44801 Bochum, Germany 2Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Nussallee 8, 53115 Bonn, Germany 3Helmholtz-Zentrum Potsdam, Deutsches GeoForschungsZentrum GFZ Sektion 3.2, Geomechanik und Rheologie, Telegrafenberg, D 429, 14473 Potsdam, Germany 4Institut für Geologische Wissenschaften, Fachrichtung Paläontologie, Freie Universität Berlin, Malteserstrasse 74–100, 12249 Berlin, Germany 5Zuse Institut Berlin, Takustrasse 7, 14195 Berlin, Germany Correspondence to: R. Hoffmann ([email protected]) Received: 28 October 2013 – Published in Biogeosciences Discuss.: 29 November 2013 Revised: 14 March 2014 – Accepted: 29 March 2014 – Published: 22 May 2014 Abstract. Several non-invasive methods are common prac- tially preserved within the surrounding rocks, requires imag- tice in natural sciences today. Here we present how they ing methods that are primarily used in non-destructive test- can be applied and contribute to current topics in cephalo- ing. The conservation of the specimen is of main importance pod (paleo-) biology. Different methods will be compared in using these methods since former techniques used destruc- terms of time necessary to acquire the data, amount of data, tive methods leading to the loss of the specimen or parts accuracy/resolution, minimum/maximum size of objects that of the specimen. -

Xenosphinctes N. Gen. (Ataxioceratidae, Lithacoceratinae), a New Rare Ammonite Genus from the Lower Tithonian (Hybonotum Zone) of SW Germany
VOLUMINA JURASSICA, 2017, XV: 155–160 DOI: 10.5604/01.3001.0010.7170 Xenosphinctes n. gen. (Ataxioceratidae, Lithacoceratinae), a new rare ammonite genus from the Lower Tithonian (Hybonotum Zone) of SW Germany Armin SCHERZINGER1, Günter SCHWEIGERT2 Key words: ammonites, Xenosphinctes berkai, Early Tithonian, biostratigraphy, Germany. Abstract. The new monotypic ammonite genus Xenosphinctes (type species: Xenosphinctes berkai n. sp.) is established. It is recorded from the Upper Jurassic, Early Tithonian, Hybonotum Zone, Riedense Subzone, eigeltingense α horizon from the Talmühle, N of Engen, Baden-Württemberg, SW Germany. INTRODUCTION AMMONITE FAUNA OF THE EIGELTINGENSE α HORIZON The Hangende-Bankkalke Formation is the youngest formation in the Upper Jurassic of Swabia. Its base is usual- The first comprehensive review of ammonites from the ly formed by an unconformity with a marked change in li- uppermost Jurassic in Swabia was provided by Berckhemer thology from coarse-grained bioclastic limestones or marl- & Hölder (1959). However, therein the focus was on the am- stones to pure mudstones (Schweigert, 1996). Its type monite fauna of the Late Kimmeridgian Beckeri Zone, and locality is an abandoned limestone quarry near the village of only very few taxa from higher levels have been reported. Eigeltingen, and its base is best exposed in a nearby section Ohmert & Zeiss (1980) started with a description of the am- along a railroad cut, NW of the Talmühle hamlet (Fig. 1). monite fauna of the Hangende-Bankkalke Formation in the The most abundant macrofossils in these basal parts of the western part of the Swabian Alb. On that occasion several formation are ammonites. Many of them are fragmented due important taxa from the basal part of this formation which to predation, but some of them are quite well-preserved as are typical of the former eigeltingense horizon were intro- steinkerns with superimposed sculpture of the outer shell. -

Correlations of the Jurassic Sediments: Infra-Getic Unit
GEOLO[KI ANALI BALKANSKOGA POLUOSTRVA 67 19–33 BEOGRAD, decembar 2006 ANNALES GÉOLOGIQUES DE LA PÉNINSULE BALKANIQUE BELGRADE, December 2006 Tran-sborder (south-east Serbia/west Bulgaria) correlations of the Jurassic sediments: Infra-Getic Unit 1 2 PLATON TCHOUMATCHENCO , DRAGOMAN RABRENOVI] , 3 4 BARBARA RADULOVI] & VLADAN RADULOVI] Abstract. The Infra-Getic Unit is a palaeogeographic unit, predestined by palaeotectonics. From the point of view of geological heritage, it represents a geosites framework. For the purpose of the correlation, the Serbian sections of Lukanja, Bogorodica Monastery, Rosoma~ and Senokos, as well as the Bulgarian sections of Komshtitsa, Gintsi, and Stanyantsi were used. The Jurassic sediments of the Infra-Getic Unit crop out on the southern slops of the Stara Planina Mountain in east Serbia and west Bulgaria. The Lower Jurassic started with continental and continental-marine sediments (clays and sandstones) (Lukanja clastics and Lukanja coal beds in Serbia and the Tuden Formation in Bulgaria) and continue with Lukanja quartz sandstones (Serbia) and the Kostina Formation (Bulgaria). These sediments are covered by Lukanja brachiopod beds and Lukanja limestones (Serbia) and the Romanov Dol, Ravna and Dolni Loukovit Members of the Ozirovo Formation (Bulgaria) pre- dominantly consist of bioclastic limestones. The sedimentations follow with Lukanja belemnites-gryphaea beds (marls and clayey limestones), which in Bulgaria correspond to the Bukorovtsi Member (also marls and clayey limestones) of the Ozirovo Formation. The Middle Jurassic sedimentation started with black shales with Bossitra alpine. These sediments are individualized in Serbia as Senokos aleurolites and clays and in Bulgaria they are known as the Etropole Formation. In Serbia the section continues with sandstones called Vodeni~ki sandstones of Bajocian age, known in Bulgaria as the Dobrogled Member of the Polaten Formation. -

Organic Carbon Isotope Chemostratigraphy of Late Jurassic Early Cretaceous Arctic Canada
University of Plymouth PEARL https://pearl.plymouth.ac.uk Faculty of Science and Engineering School of Geography, Earth and Environmental Sciences Finding the VOICE: organic carbon isotope chemostratigraphy of Late Jurassic Early Cretaceous Arctic Canada Galloway, JM http://hdl.handle.net/10026.1/15324 10.1017/s0016756819001316 Geological Magazine Cambridge University Press (CUP) All content in PEARL is protected by copyright law. Author manuscripts are made available in accordance with publisher policies. Please cite only the published version using the details provided on the item record or document. In the absence of an open licence (e.g. Creative Commons), permissions for further reuse of content should be sought from the publisher or author. Proof Delivery Form Geological Magazine Date of delivery: Journal and vol/article ref: geo 1900131 Number of pages (not including this page): 15 This proof is sent to you on behalf of Cambridge University Press. Please check the proofs carefully. Make any corrections necessary on a hardcopy and answer queries on each page of the proofs Please return the marked proof within 2 days of receipt to: [email protected] Authors are strongly advised to read these proofs thoroughly because any errors missed may appear in the final published paper. This will be your ONLY chance to correct your proof. Once published, either online or in print, no further changes can be made. To avoid delay from overseas, please send the proof by airmail or courier. If you have no corrections to make, please email [email protected] to save having to return your paper proof. If corrections are light, you can also send them by email, quoting both page and line number. -

The Geologic Time Scale Is the Eon
Exploring Geologic Time Poster Illustrated Teacher's Guide #35-1145 Paper #35-1146 Laminated Background Geologic Time Scale Basics The history of the Earth covers a vast expanse of time, so scientists divide it into smaller sections that are associ- ated with particular events that have occurred in the past.The approximate time range of each time span is shown on the poster.The largest time span of the geologic time scale is the eon. It is an indefinitely long period of time that contains at least two eras. Geologic time is divided into two eons.The more ancient eon is called the Precambrian, and the more recent is the Phanerozoic. Each eon is subdivided into smaller spans called eras.The Precambrian eon is divided from most ancient into the Hadean era, Archean era, and Proterozoic era. See Figure 1. Precambrian Eon Proterozoic Era 2500 - 550 million years ago Archaean Era 3800 - 2500 million years ago Hadean Era 4600 - 3800 million years ago Figure 1. Eras of the Precambrian Eon Single-celled and simple multicelled organisms first developed during the Precambrian eon. There are many fos- sils from this time because the sea-dwelling creatures were trapped in sediments and preserved. The Phanerozoic eon is subdivided into three eras – the Paleozoic era, Mesozoic era, and Cenozoic era. An era is often divided into several smaller time spans called periods. For example, the Paleozoic era is divided into the Cambrian, Ordovician, Silurian, Devonian, Carboniferous,and Permian periods. Paleozoic Era Permian Period 300 - 250 million years ago Carboniferous Period 350 - 300 million years ago Devonian Period 400 - 350 million years ago Silurian Period 450 - 400 million years ago Ordovician Period 500 - 450 million years ago Cambrian Period 550 - 500 million years ago Figure 2. -
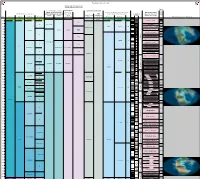
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

A Review of the Classification of Jurassic Aspidoceratid Ammonites – the Superfamily Aspidoceratoidea
VOLUMINA JURASSICA, 2020, XVIII (1): 47–52 DOI: 10.7306/VJ.18.4 A review of the classification of Jurassic aspidoceratid ammonites – the Superfamily Aspidoceratoidea Horacio PARENT1, Günter SCHWEIGERT2, Armin SCHERZINGER3 Key words: Superfamily Aspidoceratoidea, Aspidoceratidae, Epipeltoceratinae emended, Peltoceratidae, Gregoryceratinae nov. subfam. Abstract. The aspidoceratid ammonites have been traditionally included in the superfamily Perisphinctoidea. However, the basis of this is unclear for they bear unique combinations of characters unknown in typical perisphinctoids: (1) the distinct laevaptychus, (2) stout shells with high growth rate of the whorl section area, (3) prominent ornamentation with tubercles, spines and strong growth lines running in parallel over strong ribs, (4) lack of constrictions, (5) short to very short bodychamber, and (6) sexual dimorphism characterized by minia- turized microconchs and small-sized macroconchs besides the larger ones, including changes of sex during ontogeny in many cases. Considering the uniqueness of these characters we propose herein to raise the family Aspidoceratidae to the rank of a superfamily Aspi- doceratoidea, ranging from the earliest Late Callovian to the Early Berriasian Jacobi Zone. The new superfamily includes two families, Aspidoceratidae (Aspidoceratinae, Euaspidoceratinae, Epipeltoceratinae and Hybonoticeratinae), and Peltoceratidae (Peltoceratinae and Gregoryceratinae nov. subfam.). The highly differentiated features of the aspidoceratoids indicate that their life-histories -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

301083243.Pdf
AMEGHINIANA ISSN 0002-7014 Revista de la Asociación Paleontológica Argentina Tomo XII Diciembre de 1975 N?4 THE INDO-PACIFIC AMMONITE MAY AITES IN THE OXFORDIAN OF THESOUTHERN ANDES By P. N. STIPANICIC', G. E. G. WESTERMANN 2 and A. C. RJCCARDI3 ABSTRACT: Oxfordian Iitho- and biostratigraphy of the Chilean and Argentine Andes is reviewed (P. N. Stipanicic). Within the Chacay Group, the Lower to basal Upper Oxfordian La Manga Formation, below, mostly detrital and biogenic, and the Upper Oxfordian Au- quilco Formation, above, mainly chemical, are distinguished. The La Manga Formation (with Gryphaea calceola lumachelle) is rich in ammonite faunas, particularly of thc upper Cordatum to lower Canaliculatum Zones. In Neuquén and Mendoza provinces of Argentina, the Pli- catilis Zone or Middle Oxfordian has yielded Perísphinctes spp., Euaspidoceras spp., Aspido- ceras spp., together with Mayaítes (Araucanites ) stípanícfcí, M. (A.) reyesi, and M. (A.) mulai, Westermann et Riccardi subgen. et spp. nov. The first find of Mayaitidae outside the Indo-Pacific province is discussed in light of _plate-tectonic theory. RESUMEN: La revisron Iito- y bioestratigráfica del Oxfordiano de los Andes de Argentina y Chile (P. N. Stipanicic) ha permitido reconocer dentro del Grupo Chacay: 1) abajo, la Formación La Manga, mayormente detrítica y biogénica, del Oxfordiano inferior-superior basa!, y 2) arriba, la Formación Auquilco, mayormente química, del Oxfordiano superior. La For- mación La Manga (con lumachelas de Gryphaea calceola) contiene abundante cantidad de amonitas, particularmente de las Zonas de Cordatum superior a Canaliculatum inferior. En las provincias de Mendoza y Neuquén,Argentina, la Zona de Plicatilis (Oxfordiano medio) contiene Perispbinctes spp., Euaspidoceras spp., Aspidoceras spp., conjuntamente con Mayaites (Araucanites) stipanicici, M.