Liver Elimination of Bacteria Taken up by the Neutrophil
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Acute Liver Failure J G O’Grady
148 Postgrad Med J: first published as 10.1136/pgmj.2004.026005 on 4 March 2005. Downloaded from REVIEW Acute liver failure J G O’Grady ............................................................................................................................... Postgrad Med J 2005;81:148–154. doi: 10.1136/pgmj.2004.026005 Acute liver failure is a complex multisystemic illness that account for most cases, but a significant number of patients have no definable cause and are evolves quickly after a catastrophic insult to the liver classified as seronegative or of being of indeter- leading to the development of encephalopathy. The minate aetiology. Paracetamol is the commonest underlying aetiology and the pace of progression strongly cause in the UK and USA.2 Idiosyncratic reac- tions comprise another important group. influence the clinical course. The commonest causes are paracetamol, idiosyncratic drug reactions, hepatitis B, and Viral seronegative hepatitis. The optimal care is multidisciplinary ALF is an uncommon complication of viral and up to half of the cases receive liver transplants, with hepatitis, occurring in 0.2%–4% of cases depend- ing on the underlying aetiology.3 The risk is survival rates around 75%–90%. Artificial liver support lowest with hepatitis A, but it increases with the devices remain unproven in efficacy in acute liver failure. age at time of exposure. Hepatitis B can be associated with ALF through a number of ........................................................................... scenarios (table 2). The commonest are de novo infection and spontaneous surges in viral repli- cation, while the incidence of the delta virus cute liver failure (ALF) is a complex infection seems to be decreasing rapidly. multisystemic illness that evolves after a Vaccination should reduce the incidence of Acatastrophic insult to the liver manifesting hepatitis A and B, while antiviral drugs should in the development of a coagulopathy and ameliorate replication of hepatitis B. -

Differential Metabolism of Alprazolam by Liver and Brain Cytochrome (P4503A) to Pharmacologically Active Metabolite
The Pharmacogenomics Journal (2002) 2, 243–258 2002 Nature Publishing Group All rights reserved 1470-269X/02 $25.00 www.nature.com/tpj ORIGINAL ARTICLE Differential metabolism of alprazolam by liver and brain cytochrome (P4503A) to pharmacologically active metabolite HV Pai1,2* ABSTRACT SC Upadhya1,2* Cytochrome P450 (P450) is a superfamily of enzymes which mediates metab- 1 olism of xenobiotics including drugs. Alprazolam, an anti-anxiety agent, is SJ Chinta * metabolized in rat and human liver by P4503A1 and P4503A4 respectively, SN Hegde1 to 4-hydroxy alprazolam (4-OHALP, pharmacologically less active) and ␣- V Ravindranath1,2 hydroxy alprazolam (␣-OHALP, pharmacologically more active). We exam- ined P450 mediated metabolism of alprazolam by rat and human brain 1Department of Neurochemistry, National microsomes and observed that the relative amount of ␣-OHALP formed in Institute of Mental Health & Neurosciences, brain was higher than liver. This biotransformation was mediated by a P450 Bangalore, India; 2National Brain Research Centre, ICGEB Campus, Aruna Asaf Ali Marg, isoform belonging to P4503A subfamily, which is constitutively expressed in New Delhi , India neuronal cells in rat and human brain. The formation of larger amounts of ␣-OHALP in neurons points to local modulation of pharmacological activity Correspondence: in brain, at the site of action of the anti-anxiety drug. Since hydroxy metab- V Ravindranath, National Brain Research olites of alprazolam are hydrophilic and not easily cleared through blood- Centre, ICGEB Campus, Aruna Asaf Ali ␣ Marg, New Delhi - 110 067, India CSF barrier, -OHALP would potentially have a longer half-life in brain. Tel: +91 124 630 8317 The Pharmacogenomics Journal (2002) 2, 243–258. -

Fact Sheet - Symptoms of Pancreatic Cancer
Fact Sheet - Symptoms of Pancreatic Cancer Diagnosis Pancreatic cancer is often difficult to diagnose, because the pancreas lies deep in the abdomen, behind the stomach, so tumors are not felt during a physical exam. Pancreatic cancer is often called the “silent” cancer because the tumor can grow for many years before it causes pressure, pain, or other signs of illness. When symptoms do appear, they can vary depending on the size of the tumor and where it is located on the pancreas. For these reasons, the symptoms of pancreatic cancer are seldom recognized until the cancer has progressed to an advanced stage and often spread to other areas of the body. General Symptoms Pain The first symptom of pancreatic cancer is often pain, because the tumors invade nerve clusters. Pain can be felt in the stomach area and/or in the back. The pain is generally worse after eating and when lying down, and is sometimes relieved by bending forward. Pain is more common in cancers of the body and tail of the pancreas. The abdomen may also be generally tender or painful if the liver, pancreas or gall bladder are inflamed or enlarged. It is important to keep in mind that there are many other causes of abdominal and back pain! Jaundice More than half of pancreatic cancer sufferers have jaundice, a yellowing of the skin and whites of the eyes. Jaundice is caused by a build-up bilirubin, a substance which is made in the liver and a component of bile. Bilirubin contains a lot of yellow pigment, and gives bile it’s color. -

Hepatitis B? HEPATITIS B Hepatitis B Is a Contagious Liver Disease That Results from Infection with the Hepatitis B Virus
What is Hepatitis B? HEPATITIS B Hepatitis B is a contagious liver disease that results from infection with the Hepatitis B virus. When first infected, a person can develop Are you at risk? an “acute” infection, which can range in severity from a very mild illness with few or no symptoms to a serious condition requiring hospitalization. Acute Hepatitis B refers to the first 6 months after someone is exposed to the Hepatitis B virus. Some people are able to fight the infection and clear the virus. For others, the infection remains and leads to a “chronic,” or lifelong, illness. Chronic Hepatitis B refers to the illness that occurs when the Hepatitis B virus remains in a person’s body. Over time, the infection can cause serious health problems. How is Hepatitis B spread? Hepatitis B is usually spread when blood, semen, or other body fluids from a person infected with the Hepatitis B virus enter the body of someone who is not infected. This can happen through having sex with an infected partner; sharing needles, syringes, or other injection drug equipment; or from direct contact with the blood or open sores of an infected person. Hepatitis B can also be passed from an infected mother to her baby at birth. Who should be tested for Hepatitis B? Approximately 1.2 million people in the United States and 350 million people worldwide have Hepatitis B. Testing for Hepatitis B is recommended for certain groups of people, including: Most are unaware of their infection. ■ People born in Asia, Africa, and other regions with moderate or high rates Is Hepatitis B common? of Hepatitis B (see map) Yes. -
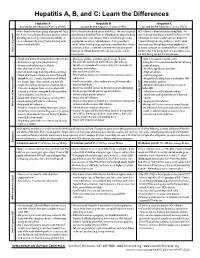
Hepatitis A, B, and C: Learn the Differences
Hepatitis A, B, and C: Learn the Differences Hepatitis A Hepatitis B Hepatitis C caused by the hepatitis A virus (HAV) caused by the hepatitis B virus (HBV) caused by the hepatitis C virus (HCV) HAV is found in the feces (poop) of people with hepa- HBV is found in blood and certain body fluids. The virus is spread HCV is found in blood and certain body fluids. The titis A and is usually spread by close personal contact when blood or body fluid from an infected person enters the body virus is spread when blood or body fluid from an HCV- (including sex or living in the same household). It of a person who is not immune. HBV is spread through having infected person enters another person’s body. HCV can also be spread by eating food or drinking water unprotected sex with an infected person, sharing needles or is spread through sharing needles or “works” when contaminated with HAV. “works” when shooting drugs, exposure to needlesticks or sharps shooting drugs, through exposure to needlesticks on the job, or from an infected mother to her baby during birth. or sharps on the job, or sometimes from an infected How is it spread? Exposure to infected blood in ANY situation can be a risk for mother to her baby during birth. It is possible to trans- transmission. mit HCV during sex, but it is not common. • People who wish to be protected from HAV infection • All infants, children, and teens ages 0 through 18 years There is no vaccine to prevent HCV. -

How Is Alcohol Metabolized by the Body?
Overview: How Is Alcohol Metabolized by the Body? Samir Zakhari, Ph.D. Alcohol is eliminated from the body by various metabolic mechanisms. The primary enzymes involved are aldehyde dehydrogenase (ALDH), alcohol dehydrogenase (ADH), cytochrome P450 (CYP2E1), and catalase. Variations in the genes for these enzymes have been found to influence alcohol consumption, alcohol-related tissue damage, and alcohol dependence. The consequences of alcohol metabolism include oxygen deficits (i.e., hypoxia) in the liver; interaction between alcohol metabolism byproducts and other cell components, resulting in the formation of harmful compounds (i.e., adducts); formation of highly reactive oxygen-containing molecules (i.e., reactive oxygen species [ROS]) that can damage other cell components; changes in the ratio of NADH to NAD+ (i.e., the cell’s redox state); tissue damage; fetal damage; impairment of other metabolic processes; cancer; and medication interactions. Several issues related to alcohol metabolism require further research. KEY WORDS: Ethanol-to acetaldehyde metabolism; alcohol dehydrogenase (ADH); aldehyde dehydrogenase (ALDH); acetaldehyde; acetate; cytochrome P450 2E1 (CYP2E1); catalase; reactive oxygen species (ROS); blood alcohol concentration (BAC); liver; stomach; brain; fetal alcohol effects; genetics and heredity; ethnic group; hypoxia The alcohol elimination rate varies state of liver cells. Chronic alcohol con- he effects of alcohol (i.e., ethanol) widely (i.e., three-fold) among individ- sumption and alcohol metabolism are on various tissues depend on its uals and is influenced by factors such as strongly linked to several pathological concentration in the blood T chronic alcohol consumption, diet, age, consequences and tissue damage. (blood alcohol concentration [BAC]) smoking, and time of day (Bennion and Understanding the balance of alcohol’s over time. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

Hepatitis a Is a Serious Liver Disease...Vaccination Can Protect You!
Hepatitis A Is a Serious Liver Disease vaccination can protect you! What is hepatitis A? Who should get hepatitis A vaccine? Hepatitis A is a serious liver disease caused by infection with The following groups of people should get vaccinated the hepatitis A virus. against hepatitis A virus if they haven’t already been vaccinated: How is hepatitis A virus spread? • Any person who wishes to be protected from hepatitis A Hepatitis A virus is usually spread from getting particles virus infection of fecal material (poop) into your mouth that are too small • All children who are age 12 through 23 months to be seen. This can happen through household or sexual • All unvaccinated children age 2 through 18 years contact with an infected person or by eating hepatitis A • Men who have sex with men virus-contaminated food, using virus-contaminated street • Users of street drugs (injecting and non-injecting) drugs, or by drinking hepatitis A virus-contaminated water. • People who are homeless or in temporary housing (such Casual contact, such as in a school or work setting, has not as a shelter) been known to spread hepatitis A virus. • People age 6 months and older who travel or work in any area of the world except the U.S., Canada, parts of West- What are the symptoms of hepatitis A virus infection? ern Europe, Japan, New Zealand, and Australia • People who will have close personal contact with an Infected people can have no symptoms at all or be extremely international adoptee from a country where hepatitis A ill. -

GLOSSARYGLOSSARY Medical Terms Common to Hepatology
GLOSSARYGLOSSARY Medical Terms Common to Hepatology Abdomen (AB-doh-men): The area between the chest and the hips. Contains the stomach, small intestine, large intestine, liver, gallbladder, pancreas and spleen. Absorption (ub-SORP-shun): The way nutrients from food move from the small intestine into the cells in the body. Acetaminophen (uh-seat-uh-MIN-oh-fin): An active ingredient in some over-the-counter fever reducers and pain relievers, including Tylenol. Acute (uh-CUTE): A disorder that has a sudden onset. Alagille Syndrome (al-uh-GEEL sin-drohm): A condition when the liver has less than the normal number of bile ducts. It is associated with other characteristics such as particular facies, abnormal pulmonary artery and abnormal vertebral bodies. Alanine Aminotransferase or ALT (AL-ah-neen uh-meen-oh-TRANZ-fur-ayz): An enzyme produced by hepatocytes, the major cell types in the liver. As cells are damaged, ALT leaks out into the bloodstream. ALT levels above normal may indicate liver damage. Albumin (al-BYEW-min): A protein that is synthesized by the liver and secreted into the blood. Low levels of albumin in the blood may indicate poor liver function. Alimentary Canal (al-uh-MEN-tree kuh-NAL): See Gastrointestinal (GI) Tract. Alkaline Phosphatase (AL-kuh-leen FOSS-fuh-tayz): Proteins or enzymes produced by the liver when bile ducts are blocked. Allergy (AL-ur-jee): A condition in which the body is not able to tolerate or has a reaction to certain foods, animals, plants, or other substances. Amino Acids (uh-MEE-noh ASS-udz): The basic building blocks of proteins. -

COVID-19 and Liver Cirrhosis Important Information for Patients and Their Families
COVID-19 and Liver Cirrhosis Important Information for Patients and Their Families The American Association for the Study of Liver Diseases (AASLD) is committed to helping you understand coronavirus disease 2019 (COVID-19) infection and prevention in people with liver cirrhosis. What We Know Our understanding of COVID-19 in people with liver cirrhosis is evolving. When making decisions related to COVID-19 infections or prevention, having up-to-date information is critical. • Symptoms of COVID-19 infection include any of the following: fever, chills, drowsiness, cough, congestion or runny nose, difficulty breathing, fatigue, body aches, headache, sore throat, abdominal pain, nausea, vomiting, diarrhea, and loss of sense of taste or smell. • People with underlying cirrhosis of the liver are at a higher risk of developing severe COVID-19 illness and/or more problems from their existing liver disease if they get a COVID-19 infection, with prolonged hospitalization and increased mortality. These patients need to take careful precautions to avoid COVID-19 infection. COVID-19 may affect the processes and procedures for screening, diagnosis, and treatment of liver cirrhosis. • Cirrhosis, or scarring of the liver, can be caused by many chronic liver diseases, including viral hepatitis, as well as excessive alcohol intake, obesity, diabetes, diseases of the bile ducts, and a variety of toxic, metabolic, or other inherited diseases. • Most people with liver disease are asymptomatic. Complications, such as yellowing of the skin and eyes from jaundice, internal bleeding (varices), mental confusion (hepatic encephalopathy), and/or swollen belly from ascites, may take years to develop, so patients are often unaware of the severity of their condition and the slow, progressive damage. -

Kupffer Cell Release of Platelet Activating Factor Drives Dose Limiting Toxicities of Nucleic Acid Nanocarriers
bioRxiv preprint doi: https://doi.org/10.1101/2020.02.11.944504; this version posted February 12, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC 4.0 International license. Kupffer Cell Release of Platelet Activating Factor Drives Dose Limiting Toxicities of Nucleic Acid Nanocarriers. Meredith A. Jackson1, Shrusti S. Patel1, Fang Yu1, Matthew A. Cottam2, Evan B. Glass1, Bryan R. Dollinger1, Ella N. Hoogenboezem1, Prarthana Patil1, Danielle D. Liu1, Isom B. Kelly1, Sean K. Bedingfield1, Allyson R. King1, Rachel E. Miles1, Alyssa M. Hasty2,3, Todd D. Giorgio1, Craig L. Duvall1*. 1Department of Biomedical Engineering, Vanderbilt University, Nashville, TN, 37235, USA 2Department of Molecular Physiology and Biophysics, Vanderbilt University School of Medicine, Nashville, TN, 37232, USA 3Veterans Affairs Tennessee Valley Healthcare System, Nashville, TN, 37212, USA *Email: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/2020.02.11.944504; this version posted February 12, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC 4.0 International license. Abstract In vivo nanocarrier-associated toxicity is a significant and poorly understood hurdle to clinical translation of siRNA nanomedicines. In this work, we demonstrate that platelet activating factor (PAF), an inflammatory lipid mediator, plays a key role in nanocarrier- associated toxicities, and that prophylactic inhibition of the PAF receptor (PAFR) completely prevents these toxicities. -

Role of Sialic Acid in Survival of Erythrocytes in the Circulation
Proc. Nati. Acad. Sci. USA Vol. 74, No. 4, pp. 1521-1524, April 1977 Biochemistry Role of sialic acid in survival of erythrocytes in the circulation: Interaction of neuraminidase-treated and untreated erythrocytes with spleen and liver at the cellular level (agglutination/erythrocyte aging/Kupffer cells/neuraminic acids/reticuloendothelial system) DAVID AMINOFF, WILLIAM F. VORDER BRUEGGE, WILLIAM C. BELL, KEITH SARPOLIS, AND REVIUS WILLIAMS Departments of Internal Medicine (Simpson Memorial Institute) and Biological Chemistry, University of Michigan, Ann Arbor, Michigan 48109 Communicated by J. L. Oncley, February 7,1977 ABSTRACT Sialidase (neuraminidase; acylneuraminyl either Sigma Biochemicals (No. C-0130, lot 15C-0037) or hydrolase; EC 3.2.1.18)treated erythrocytes obtained from Worthington Biochemicals (Class III, lot 46D084). Elemental different species are susceptible to rapid elimination from the iron particles 3-4 in size were obtained from the G.A.F. circulation and are sequestered in the liver and spleen. The /Am present studies were concerned with the mechanism of this Corp. ("carbonyl iron, S-F special"). Bovine serum albumin in clearance and how it may relate to the normal physiological crystalline form was obtained from Pentex Biochemicals (lot process of removing senescent erythrocytes from the circulation. 18). Ficoll-Paque lymphocyte isolation medium was obtained The results obtained indicate a preferential recognition of si- from Pharmacia Fine Chemicals (lot C5PO01). alidase-treated as compared to normal erythrocytes by mono- Enzymatic Treatment of Erythrocytes. Blood was collected nuclear spleen cells and Kupffer cells of the liver. This recog- in EDTA (1.4 mg/ml) from 200- to 250-g male rats (Sprague- nition manifests itself in both autologous and homologous sys- tems by adhesion of the complementary cells in the form of ro- Dawley strain) maintained on standard laboratory chow and settes, and as such could explain the removal of enzyme-treated tap water ad lib.