The Role of Comets in Panspermia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Modelling Panspermia in the TRAPPIST-1 System
October 13, 2017 Modelling panspermia in the TRAPPIST-1 system James A. Blake1,2*, David J. Armstrong1,2, Dimitri Veras1,2 Abstract The recent ground-breaking discovery of seven temperate planets within the TRAPPIST-1 system has been hailed as a milestone in the development of exoplanetary science. Centred on an ultra-cool dwarf star, the planets all orbit within a sixth of the distance from Mercury to the Sun. This remarkably compact nature makes the system an ideal testbed for the modelling of rapid lithopanspermia, the idea that micro-organisms can be distributed throughout the Universe via fragments of rock ejected during a meteoric impact event. We perform N-body simulations to investigate the timescale and success-rate of lithopanspermia within TRAPPIST-1. In each simulation, test particles are ejected from one of the three planets thought to lie within the so-called ‘habitable zone’ of the star into a range of allowed orbits, constrained by the ejection velocity and coplanarity of the case in question. The irradiance received by the test particles is tracked throughout the simulation, allowing the overall radiant exposure to be calculated for each one at the close of its journey. A simultaneous in-depth review of space microbiological literature has enabled inferences to be made regarding the potential survivability of lithopanspermia in compact exoplanetary systems. 1Department of Physics, University of Warwick, Coventry, CV4 7AL 2Centre for Exoplanets and Habitability, University of Warwick, Coventry, CV4 7AL *Corresponding author: [email protected] Contents Universe, and can propagate from one location to another. This interpretation owes itself predominantly to the works of William 1 Introduction1 Thompson (Lord Kelvin) and Hermann von Helmholtz in the 1.1 Mechanisms for panspermia...............2 latter half of the 19th Century. -
USGS Open-File Report 2005-1190, Appendix A
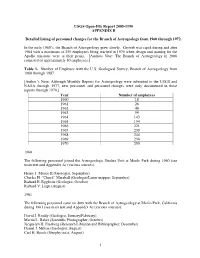
USGS Open-File Report 2005-1190 APPENDIX B Detailed listing of personnel changes for the Branch of Astrogeology from 1960 through 1972. In the early 1960’s, the Branch of Astrogeology grew slowly. Growth was rapid during and after 1964 with a maximum of 250 employees being reached in 1970 when design and training for the Apollo missions were at their peaks. [Authors Note: The Branch of Astrogeology in 2006 consisted of approximately 80 employees.] Table 1. Number of Employee with the U.S. Geological Survey, Branch of Astrogeology from 1960 through 1987 [Author’s Note: Although Monthly Reports for Astrogeology were submitted to the USGS and NASA through 1977, new personnel and personnel changes were only documented in these reports through 1970.] Year Number of employees 1960 18 1961 26 1962 40 1963 59 1964 143 1965 154 1966 221 1967 239 1968 244 1969 234 1970 250 1960 The following personnel joined the Astrogeologic Studies Unit at Menlo Park during 1960 (see main text and Appendix A) (various sources): Henry J. Moore II (Geologist, September) Charles H. “Chuck” Marshall (Geologist/Lunar mapper; September) Richard E. Eggleton (Geologist, October) Richard V. Lugn (August) 1961 The following personnel came on duty with the Branch of Astrogeology at Menlo Park, California during 1961 (see main text and Appendix A) (various sources): David J. Roddy (Geologist; January/February) Martin L. Baker (Scientific Photographer; October) Jacquelyn H. Freeberg (Research Librarian and Bibliographer; December) Daniel J. Milton (Geologist; August) Carl H. Roach (Geophysicist; August) 1 Maxine Burgess (Secretary) 1962 The following was taken from the Branch of Astrogeology Monthly Report for April 1962 from Chief, Branch of Astrogeology to V.E. -

The Primordial Earth: Hadean and Archean Eons
5th International Symposium on Strong Electromagnetic Fields and Neutron Stars 10 –13 of May, 2017 -Varadero, Cuba HABITABILITY OF THE MILKY WAY REVISITED Rolando Cárdenas and Rosmery Nodarse-Zulueta e-mail: [email protected] Planetary Science Laboratory Universidad Central “Marta Abreu” de Las Villas, Santa Clara, Cuba Abstract • The discoveries of the last three decades on deep sea and deep crust of planet Earth show that life can thrive in many places where solar radiation does not reach, using chemosynthesis instead of photosynthesis for primary production. • Underground life is relatively well protected from hazardous ionizing cosmic radiation, so above mentioned discoveries reopen the habitability budget of the Milky Way, turning potentially habitable even planetary bodies without atmosphere. • Considering this, in this work the habitability potential of the Milky Way is reconsidered. Energy Sources for Primary Habitability: - Photosynthesis: Electromagnetic Waves, mostly in the range 400-700 nm. Dominant in planetary surface. - Chemosynthesis: Energy released by redox chemical reactions. Dominant in deep sea and crust. Circumstellar Habitable Zone https://en.wikipedia.org/wiki/Circumstellar_habitable_zone. Accessed on 2017.04.28 Liquid water at surface: biased towards surface (photosynthetic) life Chemosynthesis: More common than previously thought… - Any redox process giving at least 20 kJ/mol of free energy can support microbial metabolism. The following gives 794 kJ/mol: Pohlman, J.: The biogeochemistry of anchialine caves: -

Cometary Panspermia a Radical Theory of Life’S Cosmic Origin and Evolution …And Over 450 Articles, ~ 60 in Nature
35 books: Cosmic origins of life 1976-2020 Physical Sciences︱ Chandra Wickramasinghe Cometary panspermia A radical theory of life’s cosmic origin and evolution …And over 450 articles, ~ 60 in Nature he combined efforts of generations supporting panspermia continues to Prof Wickramasinghe argues that the seeds of all life (bacteria and viruses) Panspermia has been around may have arrived on Earth from space, and may indeed still be raining down some 100 years since the term of experts in multiple fields, accumulate (Wickramasinghe et al., 2018, to affect life on Earth today, a concept known as cometary panspermia. ‘primordial soup’, referring to Tincluding evolutionary biology, 2019; Steele et al., 2018). the primitive ocean of organic paleontology and geology, have painted material not-yet-assembled a fairly good, if far-from-complete, picture COMETARY PANSPERMIA – cultural conceptions of life dating back galactic wanderers are normal features have argued that these could not into living organisms, was first of how the first life on Earth progressed A SOLUTION? to the ideas of Aristotle, and that this of the cosmos. Comets are known to have been lofted from the Earth to a coined. The question of how from simple organisms to what we can The word ‘panspermia’ comes from the may be the source of some of the have significant water content as well height of 400km by any known process. life’s molecular building blocks see today. However, there is a crucial ancient Greek roots ‘sperma’ meaning more hostile resistance the idea of as organics, and their cores, kept warm Bacteria have also been found high in spontaneously assembled gap in mainstream understanding - seed, and ‘pan’, meaning all. -

SIDE GROUP ADDITION to the POLYCYCLIC AROMATIC HYDROCARBON CORONENE by PROTON IRRADIATION in COSMIC ICE ANALOGS Max P
The Astrophysical Journal, 582:L25–L29, 2003 January 1 ᭧ 2003. The American Astronomical Society. All rights reserved. Printed in U.S.A. SIDE GROUP ADDITION TO THE POLYCYCLIC AROMATIC HYDROCARBON CORONENE BY PROTON IRRADIATION IN COSMIC ICE ANALOGS Max P. Bernstein,1,2 Marla H. Moore,3 Jamie E. Elsila,4 Scott A. Sandford,2 Louis J. Allamandola,2 and Richard N. Zare4 Received 2002 July 23; accepted 2002 November 5; published 2002 December 6 ABSTRACT ∼ Ices at 15 K consisting of the polycyclic aromatic hydrocarbon coronene (C24H12) condensed either with H2O, CO2, or CO in the ratio of 1 : 100 or greater have been subjected to MeV proton bombardment from a Van de Graaff generator. The resulting reaction products have been examined by infrared transmission- reflection-transmission spectroscopy and by microprobe laser-desorption laser-ionization mass spectrometry. Just as in the case of UV photolysis, oxygen atoms are added to coronene, yielding, in the case of H2O ices, the addition of one or more alcohol (i OH) and ketone (1CuO) side chains to the coronene scaffolding. There are, however, significant differences between the products formed by proton irradiation and the products formed by UV photolysis of coronene containing CO and CO2 ices. The formation of a coronene carboxylic i acid ( COOH) by proton irradiation is facile in solid CO but not in CO2, the reverse of what was previously observed for UV photolysis under otherwise identical conditions. This work presents evidence that cosmic- ray irradiation of interstellar or cometary ices should have contributed to the formation of aromatics bearing ketone and carboxylic acid functional groups in primitive meteorites and interplanetary dust particles. -

Habitability on Local, Galactic and Cosmological Scales
Habitability on local, Galactic and cosmological scales Luigi Secco1 • Marco Fecchio1 • Francesco Marzari1 Abstract The aim of this paper is to underline con- detectable studying our site. The Climatic Astronom- ditions necessary for the emergence and development ical Theory is introduced in sect.5 in order to define of life. They are placed at local planetary scale, at the circumsolar habitable zone (HZ) (sect.6) while the Galactic scale and within the cosmological evolution, translation from Solar to extra-Solar systems leads to a as pointed out by the Anthropic Cosmological Princi- generalized circumstellar habitable zone (CHZ) defined ple. We will consider the circumstellar habitable zone in sect.7 with some exemplifications to the Gliese-667C (CHZ) for planetary systems and a Galactic Habitable and the TRAPPIST-1 systems; some general remarks Zone (GHZ) including also a set of strong cosmologi- follow (sect.8). A first conclusion related to CHZ is cal constraints to allow life (cosmological habitability done moving toward GHZ and COSH (sect.9). The (COSH)). Some requirements are specific of a single conditions for the development of life are indeed only scale and its related physical phenomena, while others partially connected to the local scale in which a planet are due to the conspired effects occurring at more than is located. A strong interplay between different scales one scale. The scenario emerging from this analysis is exists and each single contribution to life from individ- that all the habitability conditions here detailed must ual scales is difficult to be isolated. However we will at least be met. -

The Large Scale Universe As a Quasi Quantum White Hole
International Astronomy and Astrophysics Research Journal 3(1): 22-42, 2021; Article no.IAARJ.66092 The Large Scale Universe as a Quasi Quantum White Hole U. V. S. Seshavatharam1*, Eugene Terry Tatum2 and S. Lakshminarayana3 1Honorary Faculty, I-SERVE, Survey no-42, Hitech city, Hyderabad-84,Telangana, India. 2760 Campbell Ln. Ste 106 #161, Bowling Green, KY, USA. 3Department of Nuclear Physics, Andhra University, Visakhapatnam-03, AP, India. Authors’ contributions This work was carried out in collaboration among all authors. Author UVSS designed the study, performed the statistical analysis, wrote the protocol, and wrote the first draft of the manuscript. Authors ETT and SL managed the analyses of the study. All authors read and approved the final manuscript. Article Information Editor(s): (1) Dr. David Garrison, University of Houston-Clear Lake, USA. (2) Professor. Hadia Hassan Selim, National Research Institute of Astronomy and Geophysics, Egypt. Reviewers: (1) Abhishek Kumar Singh, Magadh University, India. (2) Mohsen Lutephy, Azad Islamic university (IAU), Iran. (3) Sie Long Kek, Universiti Tun Hussein Onn Malaysia, Malaysia. (4) N.V.Krishna Prasad, GITAM University, India. (5) Maryam Roushan, University of Mazandaran, Iran. Complete Peer review History: http://www.sdiarticle4.com/review-history/66092 Received 17 January 2021 Original Research Article Accepted 23 March 2021 Published 01 April 2021 ABSTRACT We emphasize the point that, standard model of cosmology is basically a model of classical general relativity and it seems inevitable to have a revision with reference to quantum model of cosmology. Utmost important point to be noted is that, ‘Spin’ is a basic property of quantum mechanics and ‘rotation’ is a very common experience. -

Exploring Biogenic Dispersion Inside Star Clusters with System Dynamics Modeling
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 12 October 2020 doi:10.20944/preprints202010.0232.v1 Exploring Biogenic Dispersion Inside Star Clusters with System Dynamics Modeling Authors Javier D. Burgos. Corporación para la Investigacion e Innovación -CIINAS. Contact: [email protected] Diana C. Sierra. Corporación para la Investigacion e Innovación -CIINAS; Bogotá, Colombia. Contact: [email protected] Abstract The discovery of a growing number of exoplanets and even extrasolar systems supports the scientific consensus that it is possible to find other signs of life in the universe. The present work proposes for the first time, an explicit mechanism inspired by the dynamics of biological dispersion, widely used in ecology and epidemiology, to study the dispersion of biogenic units, interpreted as complex organic molecules, between rocky or water exoplanets (habitats) located inside star clusters. The 3 results of the dynamic simulation suggest that for clusters with populations lower than 4 M/ly it is not possible to obtain biogenic worlds after 5 Gyr. Above this population size, biogenic dispersion seems to follow a power law, the larger the density of worlds lesser will be the impact rate (훽.) value to obtain at least one viable biogenic Carrier habitat after 5 Gyr. Finally, when we investigate scenarios by varying β, a well-defined set of density intervals can be defined in accordance to its characteristic β value, suggesting that biogenic dispersion has a behavior of “minimal infective dose” of “minimal biogenic effective” events by interval i.e. once this dose has been achieved, doesn’t matter if additional biogenic impact events occur on the habitat. -

First Life in Primordial-Planet Oceans: the Biological Big Bang Gibson/Wickramasinghe/Schild First Life in Primordial-Planet Oceans: the Biological Big Bang
Journal of Cosmology First life in primordial-planet oceans: the biological big bang Gibson/Wickramasinghe/Schild First life in primordial-planet oceans: the biological big bang Carl H. Gibson 1,2 1 University of California San Diego, La Jolla, CA 92093-0411, USA [email protected], http://sdcc3.ucsd.edu/~ir118 N. Chandra Wickramasinghe3,4 3Cardiff Centre for Astrobiology, 24 Llwynypia Road, Lisvane, Cardiff CF14 0SY [email protected] Rudolph E. Schild5,6 5 Center for Astrophysics, 60 Garden Street, Cambridge, MA 02138, USA 6 [email protected] Abstract: A scenario is presented for the formation of first life in the universe based on hydro- gravitational-dynamics (HGD) cosmology. From HGD, the dark matter of galaxies is H-He gas dominated planets (primordial-fog-particle PFPs) in million solar mass clumps (protoglobularstar- cluster PGCs), which formed at the plasma to gas transition temperature 3000 K. Stars result from mergers of these hot-gas-planets. Over-accretion causes stars to explode as supernovae that scatter life-chemicals (C, N, O, P, S, Ca, Fe etc.) to other planets in PGC clumps and beyond. These chemicals were first collected gravitationally by merging PFPs to form H-saturated, high-pressure, dense oceans of critical-temperature 647 K water over iron-nickel cores at ~ 2 Myr. Stardust fertil- izes the formation of first life in a cosmic hot-ocean soup kitchen comprised of all planets and their moons in meteoric communication, > 10100 kg in total. Ocean freezing at 273 K slows this biologi- cal big bang at ~ 8 Myr. HGD cosmology confirms that the evolving seeds of life are scattered on intergalactic scales by Hoyle-Wickramasinghe cometary panspermia. -

The British Astronomical Association Handbook 2017
THE HANDBOOK OF THE BRITISH ASTRONOMICAL ASSOCIATION 2017 2016 October ISSN 0068–130–X CONTENTS PREFACE . 2 HIGHLIGHTS FOR 2017 . 3 CALENDAR 2017 . 4 SKY DIARY . .. 5-6 SUN . 7-9 ECLIPSES . 10-15 APPEARANCE OF PLANETS . 16 VISIBILITY OF PLANETS . 17 RISING AND SETTING OF THE PLANETS IN LATITUDES 52°N AND 35°S . 18-19 PLANETS – EXPLANATION OF TABLES . 20 ELEMENTS OF PLANETARY ORBITS . 21 MERCURY . 22-23 VENUS . 24 EARTH . 25 MOON . 25 LUNAR LIBRATION . 26 MOONRISE AND MOONSET . 27-31 SUN’S SELENOGRAPHIC COLONGITUDE . 32 LUNAR OCCULTATIONS . 33-39 GRAZING LUNAR OCCULTATIONS . 40-41 MARS . 42-43 ASTEROIDS . 44 ASTEROID EPHEMERIDES . 45-50 ASTEROID OCCULTATIONS .. ... 51-53 ASTEROIDS: FAVOURABLE OBSERVING OPPORTUNITIES . 54-56 NEO CLOSE APPROACHES TO EARTH . 57 JUPITER . .. 58-62 SATELLITES OF JUPITER . .. 62-66 JUPITER ECLIPSES, OCCULTATIONS AND TRANSITS . 67-76 SATURN . 77-80 SATELLITES OF SATURN . 81-84 URANUS . 85 NEPTUNE . 86 TRANS–NEPTUNIAN & SCATTERED-DISK OBJECTS . 87 DWARF PLANETS . 88-91 COMETS . 92-96 METEOR DIARY . 97-99 VARIABLE STARS (RZ Cassiopeiae; Algol; λ Tauri) . 100-101 MIRA STARS . 102 VARIABLE STAR OF THE YEAR (T Cassiopeiæ) . .. 103-105 EPHEMERIDES OF VISUAL BINARY STARS . 106-107 BRIGHT STARS . 108 ACTIVE GALAXIES . 109 TIME . 110-111 ASTRONOMICAL AND PHYSICAL CONSTANTS . 112-113 INTERNET RESOURCES . 114-115 GREEK ALPHABET . 115 ACKNOWLEDGEMENTS / ERRATA . 116 Front Cover: Northern Lights - taken from Mount Storsteinen, near Tromsø, on 2007 February 14. A great effort taking a 13 second exposure in a wind chill of -21C (Pete Lawrence) British Astronomical Association HANDBOOK FOR 2017 NINETY–SIXTH YEAR OF PUBLICATION BURLINGTON HOUSE, PICCADILLY, LONDON, W1J 0DU Telephone 020 7734 4145 PREFACE Welcome to the 96th Handbook of the British Astronomical Association. -

Observations of the Long-Lasting Activity of the Distant Comets 29P Schwassmann-Wachmann 1, C/2003 WT42 (LINEAR) and C/2002
Observations of the long-lasting activity of the distant comets 29P Schwassmann-Wachmann 1, C/2003 WT42 (LINEAR) and C/2002 VQ94 (LINEAR). Oleksandra V. Ivanova*a, Yury V. Skorovb,c, Pavlo P. Korsuna, Viktor L. Afanasievd and Jürgen Blumb a – Main Astronomical Observatory of NAS of Ukraine, Akademika Zabolotnoho 27, 03680 Kyiv, Ukraine * Corresponding Author. E-mail address: [email protected] b – University of Braunschweig. Institute for Geophysics and Extraterrestrial Physics Mendelssohn-Str. 3, D-38106 Braunschweig, Germany c – Max Planck Institute for Solar System Research, Max-Planck-Str. 2, D- 37191,Katlenburg-Lindau, Germany d – Special Astrophysical Observatory of the Russian AS, Nizhnij Arkhyz, 369167, Russia 1 Proposed Running Head: Observations of the long-lasting activity of the distant comets 29P Schwassmann-Wachmann 1, C/2003 WT42 (LINEAR) and C/2002 VQ94 (LINEAR). Editorial correspondence to: Dr. Oleksandra Ivanova Main Astronomical Observatory of NAS of Ukraine Akademika Zabolotnoho str. 27 03680 Kyiv Ukraine Phone: +380 44 526-47-69 E-Mail address: [email protected] 2 ABSTRACT We investigated three comets, which are active at large heliocentric distances, using observations obtained at the 6-m BTA telescope (SAO RAS, Russia) in the photometric mode of the focal reducer SCORPIO. The three comets, 29P/Schwassmann–Wachmann 1, C/2003 WT42 (LINEAR) and C/2002 VQ94 (LINEAR), were observed after their perihelion passages at heliocentric distances between 5.5 and 7.08 AU. The dust production rates in terms of Af was measured for these comets. Using these retrieved values, an average dust production rate was derived under different model assumptions. -

The Minor Planet Bulletin 44 (2017) 142
THE MINOR PLANET BULLETIN OF THE MINOR PLANETS SECTION OF THE BULLETIN ASSOCIATION OF LUNAR AND PLANETARY OBSERVERS VOLUME 44, NUMBER 2, A.D. 2017 APRIL-JUNE 87. 319 LEONA AND 341 CALIFORNIA – Lightcurves from all sessions are then composited with no TWO VERY SLOWLY ROTATING ASTEROIDS adjustment of instrumental magnitudes. A search should be made for possible tumbling behavior. This is revealed whenever Frederick Pilcher successive rotational cycles show significant variation, and Organ Mesa Observatory (G50) quantified with simultaneous 2 period software. In addition, it is 4438 Organ Mesa Loop useful to obtain a small number of all-night sessions for each Las Cruces, NM 88011 USA object near opposition to look for possible small amplitude short [email protected] period variations. Lorenzo Franco Observations to obtain the data used in this paper were made at the Balzaretto Observatory (A81) Organ Mesa Observatory with a 0.35-meter Meade LX200 GPS Rome, ITALY Schmidt-Cassegrain (SCT) and SBIG STL-1001E CCD. Exposures were 60 seconds, unguided, with a clear filter. All Petr Pravec measurements were calibrated from CMC15 r’ values to Cousins Astronomical Institute R magnitudes for solar colored field stars. Photometric Academy of Sciences of the Czech Republic measurement is with MPO Canopus software. To reduce the Fricova 1, CZ-25165 number of points on the lightcurves and make them easier to read, Ondrejov, CZECH REPUBLIC data points on all lightcurves constructed with MPO Canopus software have been binned in sets of 3 with a maximum time (Received: 2016 Dec 20) difference of 5 minutes between points in each bin.