Impact of Myrtle Rust on Post-Fire Regeneration
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Immersive Small Group Journeys
Immersive Small Group Journeys TASMANIA • ULURU • KAKADU • NEW ZEALAND • MARGARET RIVER 2021–2022 INSPIRINGJOURNEYS.COM The immersive, vast beauty of Australia and New Zealand awaits. You’ll witness the many hues of an outback sunset blend into ochre natural wonders, hear ancient languages spoken and see stories written in the sand. Indulge in native flavours and relax in dwellings nestled within the heart of your destination. This is where your journey begins. Inspiring Journeys is proud to pay respect to the continuation of cultural practices of Aboriginal, Torres Strait Islander and Māori peoples. Outback Australia Right: Local guide Peter – Karrke Aboriginal Cultural Experience in Watarrka National Park Start Exploring • 6 Meet our vibrant makers and Start Exploring Awaiting you is a series of creators, and the local storytellers whose passion it is to share with you AUSTRALIA inspiring, life enriching the very best of these unique lands. Delight the senses with exclusive 24 Northern Territory Dreaming 10 days • MNCR culinary experiences and allow The wild majesty of the Top End and the dramatic landscapes of Australia’s Red Centre combine on this unforgettable journey. your expert guide to show you the iconic sights that promise 28 A Journey to the West 7 days • IJWA moments of awe and wonder. Unique Rottnest Island, renowned wine region Margaret River and an enriching Cape Naturaliste Indigenous cultural experience await you. momen ts. Your journey will be expertly curated 32 Outback Australia: The Colour Of Red 5 days • CRUA with no detail forgotten, ensuring Journey to the Red Centre of Australia, and uncover an ancient your adventure is seamless, stress culture that dates back thousands of years. -

TML Propagation Protocols
PROPAGATION PROTOCOLS This document is intended as a guide for Tamborine Mountain Landcare members who wish to assist our regeneration projects by growing some of the plants needed. It is a work in progress so if you have anything to add to the protocols – for example a different but successful way of propagating and growing a particular plant – then please give it to Julie Lake so she can add it to the document. The idea is that our shared knowledge and experience can become a valuable part of TML's intellectual property as well as a useful source of knowledge for members. As there are many hundreds of plants native to Tamborine Mountain, the protocols list will take a long time to complete, with growing information for each plant added alphabetically as time permits. While the list is being compiled by those members with competence in this field, any TML member with a query about propagating a particular plant can post it on the website for other me mb e r s to answer. To date, only protocols for trees and shrubs have been compiled. Vines and ferns will be added later. Fruiting times given are usual for the species but many rainforest plants flower and fruit opportunistically, according to weather and other conditions unknown to us, thus fruit can be produced at any time of year. Finally, if anyone would like a copy of the protocols, contact Julie on [email protected] and she’ll send you one. ………………….. Growing from seed This is the best method for most plants destined for regeneration projects for it is usually fast, easy and ensures genetic diversity in the regenerated landscape. -

Approved Conservation Advice for Rutidosis Heterogama (Heath Wrinklewren)
This Conservation Advice was approved by the Minister/Delegate of the Minister on: 3/07/2008. Approved Conservation Advice (s266B of the Environment Protection and Biodiversity Conservation Act 1999). Approved Conservation Advice for Rutidosis heterogama (Heath Wrinklewren) This Conservation Advice has been developed based on the best available information at the time this conservation advice was approved. Description Rutidosis heterogama, Family Asteraceae, also known as the Heath Wrinklewren or Heath Wrinklewort, is a perennial herb with decumbent (reclining to lying down) to erect stems, growing to 30 cm high (Harden, 1992; DECC, 2005a). The tiny yellow flowerheads are probably borne March to April (Leigh et al., 1984), chiefly in Autumn (Harden, 1992) or November to January. Seeds are dispersed by wind (Clarke et al., 1998) and the species appears to require soil disturbance for successful recruitment (Clarke et al., 1998). Conservation Status Heath Wrinklewren is listed as vulnerable. This species is eligible for listing as vulnerable under the Environment Protection and Biodiversity Conservation Act 1999 (Cwlth) (EPBC Act) as, prior to the commencement of the EPBC Act, it was listed as vulnerable under Schedule 1 of the Endangered Species Protection Act 1992 (Cwlth). The species is also listed as vulnerable on the Threatened Species Conservation Act 1995 (NSW). Distribution and Habitat Heath Wrinklewren is confined to the North Coast and Northern Tablelands regions of NSW. It is known from the Hunter Valley to Maclean, Wooli to Evans Head, and Torrington (Harden, 1992). It occurs within the Border Rivers–Gwydir, Hunter–Central Rivers and Northern Rivers (NSW) Natural Resource Management Regions. -

Bush Foods and Fibres
Australian Plants Society NORTH SHORE GROUP Ku-ring-gai Wildflower Garden Bush foods and fibres • Plant-based bush foods, medicines and poisons can come from nectar, flowers, fruit, leaves, bark, stems, sap and roots. • Plants provide fibres and materials for making many items including clothes, cords, musical instruments, shelters, tools, toys and weapons. • A fruit is the seed-bearing structure of a plant. • Do not eat fruits that you do not know to be safe to eat. Allergic reactions or other adverse reactions could occur. • We acknowledge the Traditional Custodians of this land and pay our respects to the Elders both past, present and future for they hold the memories, traditions, culture and hope of their people. Plants as food: many native plants must be processed before they are safe to eat. Flowers, nectar, pollen, Sugars, vitamins, honey, lerps (psyllid tents) minerals, starches, manna (e.g. Ribbon Gum proteins & other nutrients Eucalyptus viminalis exudate), gum (e.g. Acacia lerp manna decurrens) Fruit & seeds Staple foods Carbohydrates (sugars, starches, fibre), proteins, fats, vitamins Leaves, stalks, roots, apical Staple foods Carbohydrates, protein, buds minerals Plants such as daisies, lilies, orchids and vines Tubers, rhyzomes were a source of starchy tubers known as Carbohydrate, fibre, yams. The yam daisy Microseris lanceolata protein, vitamins, (Asteraceae) was widespread in inland NSW minerals and other states. The native yam Dioscorea transversa grows north from Stanwell Tops into Qld and Northern Territory and can be eaten raw or roasted as can those of Trachymene incisa. 1 Plant Description of food Other notes Acacia Wattle seed is a rich source of iron, Saponins and tannins and other essential elements. -

In China: Phylogeny, Host Range, and Pathogenicity
Persoonia 45, 2020: 101–131 ISSN (Online) 1878-9080 www.ingentaconnect.com/content/nhn/pimj RESEARCH ARTICLE https://doi.org/10.3767/persoonia.2020.45.04 Cryphonectriaceae on Myrtales in China: phylogeny, host range, and pathogenicity W. Wang1,2, G.Q. Li1, Q.L. Liu1, S.F. Chen1,2 Key words Abstract Plantation-grown Eucalyptus (Myrtaceae) and other trees residing in the Myrtales have been widely planted in southern China. These fungal pathogens include species of Cryphonectriaceae that are well-known to cause stem Eucalyptus and branch canker disease on Myrtales trees. During recent disease surveys in southern China, sporocarps with fungal pathogen typical characteristics of Cryphonectriaceae were observed on the surfaces of cankers on the stems and branches host jump of Myrtales trees. In this study, a total of 164 Cryphonectriaceae isolates were identified based on comparisons of Myrtaceae DNA sequences of the partial conserved nuclear large subunit (LSU) ribosomal DNA, internal transcribed spacer new taxa (ITS) regions including the 5.8S gene of the ribosomal DNA operon, two regions of the β-tubulin (tub2/tub1) gene, plantation forestry and the translation elongation factor 1-alpha (tef1) gene region, as well as their morphological characteristics. The results showed that eight species reside in four genera of Cryphonectriaceae occurring on the genera Eucalyptus, Melastoma (Melastomataceae), Psidium (Myrtaceae), Syzygium (Myrtaceae), and Terminalia (Combretaceae) in Myrtales. These fungal species include Chrysoporthe deuterocubensis, Celoporthe syzygii, Cel. eucalypti, Cel. guang dongensis, Cel. cerciana, a new genus and two new species, as well as one new species of Aurifilum. These new taxa are hereby described as Parvosmorbus gen. -
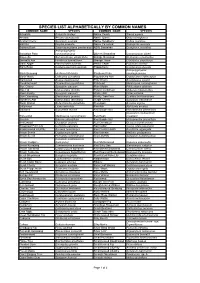
Species List Alphabetically by Common Names
SPECIES LIST ALPHABETICALLY BY COMMON NAMES COMMON NAME SPECIES COMMON NAME SPECIES Actephila Actephila lindleyi Native Peach Trema aspera Ancana Ancana stenopetala Native Quince Guioa semiglauca Austral Cherry Syzygium australe Native Raspberry Rubus rosifolius Ball Nut Floydia praealta Native Tamarind Diploglottis australis Banana Bush Tabernaemontana pandacaqui NSW Sassafras Doryphora sassafras Archontophoenix Bangalow Palm cunninghamiana Oliver's Sassafras Cinnamomum oliveri Bauerella Sarcomelicope simplicifolia Orange Boxwood Denhamia celastroides Bennetts Ash Flindersia bennettiana Orange Thorn Citriobatus pauciflorus Black Apple Planchonella australis Pencil Cedar Polyscias murrayi Black Bean Castanospermum australe Pepperberry Cryptocarya obovata Archontophoenix Black Booyong Heritiera trifoliolata Picabeen Palm cunninghamiana Black Wattle Callicoma serratifolia Pigeonberry Ash Cryptocarya erythroxylon Blackwood Acacia melanoxylon Pink Cherry Austrobuxus swainii Bleeding Heart Omalanthus populifolius Pinkheart Medicosma cunninghamii Blue Cherry Syzygium oleosum Plum Myrtle Pilidiostigma glabrum Blue Fig Elaeocarpus grandis Poison Corkwood Duboisia myoporoides Blue Lillypilly Syzygium oleosum Prickly Ash Orites excelsa Blue Quandong Elaeocarpus grandis Prickly Tree Fern Cyathea leichhardtiana Blueberry Ash Elaeocarpus reticulatus Purple Cherry Syzygium crebrinerve Blush Walnut Beilschmiedia obtusifolia Red Apple Acmena ingens Bollywood Litsea reticulata Red Ash Alphitonia excelsa Bolwarra Eupomatia laurina Red Bauple Nut Hicksbeachia -

Pre-Infection Stages of Austropuccinia Psidii in the Epidermis of Eucalyptus Hybrid Leaves with Different Resistance Levels
Article Pre-Infection Stages of Austropuccinia psidii in the Epidermis of Eucalyptus Hybrid Leaves with Different Resistance Levels Renata Ruiz Silva-Souza 1, André Costa da Silva 2, Roberto Antônio Rodella 3, José Eduardo Serrão 4, José Cola Zanuncio 5 and Edson Luiz Furtado 1,* 1 Departamento de Produção Vegetal, Faculdade de Ciências Agronômicas–UNESP, Botucatu 18610-307, São Paulo, Brasil; [email protected] 2 Departamento de Fitopatologia, Universidade Federal de Viçosa, Viçosa 36570-900, Minas Gerais, Brasil; [email protected] 3 Departamento de Botânica, Instituto de Biociências de Botucatu–UNESP, Botucatu 18618-000, São Paulo, Brasil; [email protected] 4 Departamento de Biologia Geral, Universidade Federal de Viçosa, Viçosa 36570-900, Minas Gerais, Brasil; [email protected] 5 Departamento de Entomologia/BIOAGRO, Universidade Federal de Viçosa, Viçosa 36570-900, Minas Gerais, Brasil; [email protected] * Correspondence: [email protected]; Tel.: +55-31-3899-2924 Received: 25 July 2017; Accepted: 20 September 2017; Published: 25 September 2017 Abstract: Rust is a major Eucalyptus spp. disease, which is especially damaging for early-stage plants. The aim of this study was to verify the pre-infection process of Austropuccinia psidii (A. psidii) in the leaves of three phenological stages of Eucalyptus clones with different resistance levels. Plants from the hybrids of Eucalyptus urophylla × Eucalyptus grandis (E. grandis) with variable levels of resistance to this disease were used. The pathogen was inoculated in vitro on abaxial leaf discs of first, third, and fifth leaf stages and maintained under conditions suitable for disease development. Subsequently, samples from these discs were collected 24 and 120 h after inoculation and processed using scanning electron microscopy analysis. -

Australia-15-Index.Pdf
© Lonely Planet 1091 Index Warradjan Aboriginal Cultural Adelaide 724-44, 724, 728, 731 ABBREVIATIONS Centre 848 activities 732-3 ACT Australian Capital Wigay Aboriginal Culture Park 183 accommodation 735-7 Territory Aboriginal peoples 95, 292, 489, 720, children, travel with 733-4 NSW New South Wales 810-12, 896-7, 1026 drinking 740-1 NT Northern Territory art 55, 142, 223, 823, 874-5, 1036 emergency services 725 books 489, 818 entertainment 741-3 Qld Queensland culture 45, 489, 711 festivals 734-5 SA South Australia festivals 220, 479, 814, 827, 1002 food 737-40 Tas Tasmania food 67 history 719-20 INDEX Vic Victoria history 33-6, 95, 267, 292, 489, medical services 726 WA Western Australia 660, 810-12 shopping 743 land rights 42, 810 sights 727-32 literature 50-1 tourist information 726-7 4WD 74 music 53 tours 734 hire 797-80 spirituality 45-6 travel to/from 743-4 Fraser Island 363, 369 Aboriginal rock art travel within 744 A Arnhem Land 850 walking tour 733, 733 Abercrombie Caves 215 Bulgandry Aboriginal Engraving Adelaide Hills 744-9, 745 Aboriginal cultural centres Site 162 Adelaide Oval 730 Aboriginal Art & Cultural Centre Burrup Peninsula 992 Adelaide River 838, 840-1 870 Cape York Penninsula 479 Adels Grove 435-6 Aboriginal Cultural Centre & Keep- Carnarvon National Park 390 Adnyamathanha 799 ing Place 209 Ewaninga 882 Afghan Mosque 262 Bangerang Cultural Centre 599 Flinders Ranges 797 Agnes Water 383-5 Brambuk Cultural Centre 569 Gunderbooka 257 Aileron 862 Ceduna Aboriginal Arts & Culture Kakadu 844-5, 846 air travel Centre -

Garden Escapes & Other Weeds in Bushland and Reserves a Responsible Gardening Guide for the Sydney Region
Garden Escapes & Other Weeds in Bushland and Reserves A responsible gardening guide for the Sydney Region Sydney Weeds Committees Sydney Central Sydney South West Sydney North Sydney West – Blue Mountains C O N T E N T S General Information 3 Vines & Scramblers 6 Ground Covers 20 Bulbous & Succulent Weeds 34 Grass Weeds 51 Shrub Weeds 57 Tree Weeds 64 Water Weeds 74 Help Protect Your Local Environment 77 Common Plant Parts 78 Bibliography 79 Plant Me Instead 80 Index & Acknowledments 82 Reprinted 2012- Updated in 2018 Booklet adapted and reproduced with permission of Great Lakes Council The Problem What is a weed? Plants escape from gardens in a WEEDS are plants that don’t belong variety of ways, but one main cause where they are. They can include of spread from gardens is by green plants from other countries but are also waste dumping in bushland and road sometimes from other parts of Australia. reserves. This practice is harmful to the Weeds can be harmful to human and bush for many reasons, such as: animals. They also affect the ecology and appearance of bushland areas and s introducing weeds (plant fragments, waterways. bulbs, roots, tubers, seeds, spores) Weeds often grow faster than s smothering native plants native plants and out-compete them to become dominant in natural areas. The s changing the soil and ideal growing natural pests or diseases that would conditions for native plants otherwise control their growth are lacking s increasing fi re risk by increasing as the plants have been introduced from fuel loads. somewhere else. -

The Future of World Heritage in Australia
Keeping the Outstanding Exceptional: The Future of World Heritage in Australia Editors: Penelope Figgis, Andrea Leverington, Richard Mackay, Andrew Maclean, Peter Valentine Editors: Penelope Figgis, Andrea Leverington, Richard Mackay, Andrew Maclean, Peter Valentine Published by: Australian Committee for IUCN Inc. Copyright: © 2013 Copyright in compilation and published edition: Australian Committee for IUCN Inc. Reproduction of this publication for educational or other non-commercial purposes is authorised without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Citation: Figgis, P., Leverington, A., Mackay, R., Maclean, A., Valentine, P. (eds). (2012). Keeping the Outstanding Exceptional: The Future of World Heritage in Australia. Australian Committee for IUCN, Sydney. ISBN: 978-0-9871654-2-8 Design/Layout: Pixeldust Design 21 Lilac Tree Court Beechmont, Queensland Australia 4211 Tel: +61 437 360 812 [email protected] Printed by: Finsbury Green Pty Ltd 1A South Road Thebarton, South Australia Australia 5031 Available from: Australian Committee for IUCN P.O Box 528 Sydney 2001 Tel: +61 416 364 722 [email protected] http://www.aciucn.org.au http://www.wettropics.qld.gov.au Cover photo: Two great iconic Australian World Heritage Areas - The Wet Tropics and Great Barrier Reef meet in the Daintree region of North Queensland © Photo: K. Trapnell Disclaimer: The views and opinions expressed in this publication are those of the chapter authors and do not necessarily reflect those of the editors, the Australian Committee for IUCN, the Wet Tropics Management Authority or the Australian Conservation Foundation or those of financial supporter the Commonwealth Department of Sustainability, Environment, Water, Population and Communities. -

Austropuccinia Psidii
This diagnostic protocol was adopted by the Standards Committee on behalf of the Commission on Phytosanitary Measures in August 2018. The annex is a prescriptive part of ISPM 27. ISPM 27 Diagnostic protocols for regulated pests DP 26: Austropuccinia psidii Adopted 2018; published 2018 CONTENTS 1. Pest Information ...............................................................................................................................2 2. Taxonomic Information ....................................................................................................................2 3. Detection ...........................................................................................................................................4 3.1 Signs and symptoms of infection ......................................................................................4 3.2 Sampling and sample preparation .....................................................................................5 3.3 Morphological detection ...................................................................................................5 3.4 Molecular detection ...........................................................................................................6 3.4.1 Preparation of plant material .............................................................................................6 3.4.2 Nucleic acid extraction ......................................................................................................6 3.4.3 Conventional PCR and sequencing ...................................................................................7 -

Brisbane Native Plants by Suburb
INDEX - BRISBANE SUBURBS SPECIES LIST Acacia Ridge. ...........15 Chelmer ...................14 Hamilton. .................10 Mayne. .................25 Pullenvale............... 22 Toowong ....................46 Albion .......................25 Chermside West .11 Hawthorne................. 7 McDowall. ..............6 Torwood .....................47 Alderley ....................45 Clayfield ..................14 Heathwood.... 34. Meeandah.............. 2 Queensport ............32 Trinder Park ...............32 Algester.................... 15 Coopers Plains........32 Hemmant. .................32 Merthyr .................7 Annerley ...................32 Coorparoo ................3 Hendra. .................10 Middle Park .........19 Rainworth. ..............47 Underwood. ................41 Anstead ....................17 Corinda. ..................14 Herston ....................5 Milton ...................46 Ransome. ................32 Upper Brookfield .......23 Archerfield ...............32 Highgate Hill. ........43 Mitchelton ...........45 Red Hill.................... 43 Upper Mt gravatt. .......15 Ascot. .......................36 Darra .......................33 Hill End ..................45 Moggill. .................20 Richlands ................34 Ashgrove. ................26 Deagon ....................2 Holland Park........... 3 Moorooka. ............32 River Hills................ 19 Virginia ........................31 Aspley ......................31 Doboy ......................2 Morningside. .........3 Robertson ................42 Auchenflower