Evolvability, Stabilizing Selection, and the Problem
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

An Introgression Analysis of Quantitative Trait Loci That Contribute to a Morphological Difference Between Drosophila Simulans and D
Copyight 0 1997 by the Genetics Societv of America An Introgression Analysis of Quantitative Trait Loci That Contribute to a Morphological Difference Between Drosophila simulans and D. mauritiana Cathy C. Laurie, John R. True, Jianjun Liu and John M. Mercer Department of Zoology, Duke University, Durham, North Carolina 27708 Manuscript received August 12, 1996 Accepted for publication October 29, 1996 ABSTRACT Drosophila simulans and D. maum'tiana differ markedly in morphology of the posterior lobe, a male- specific genitalic structure. Bothsize and shapeof the lobe can be quantifiedby a morphometric variable, PC1, derived from principal components and Fourier analyses. The genetic architectureof the species difference in PC1 was investigated previously by composite interval mapping, which revealed largely additive inheritance, with a minimum of eight quantitative trait loci (QTL) affecting the trait. This analysis was extended by introgression of marked segments of the maun'tiana third chromosome into a simulans background by repeated backcrossing. Thetwo types of experiment are consistentin suggesting that several QTL on the third chromosome may have effects in the range of 10-15% of the parental difference andthat all or nearly all QTL have effects in the same direction. Since the parental difference is large (30.4 environmental standard deviations), effects of this magnitude can produce alternative homozygotes with little overlap in phenotype. However, these estimates may not reflect the effects of individual loci, since each interval or introgressed segment may contain multiple QTL. The consistent direction of allelic effects suggests a history of directional selection on the posterior lobe. HE genetic analysis of a quantitative trait usually ual genes that affect continuously variable traits. -

IN EVOLUTION JACK LESTER KING UNIVERSITY of CALIFORNIA, SANTA BARBARA This Paper Is Dedicated to Retiring University of California Professors Curt Stern and Everett R
THE ROLE OF MUTATION IN EVOLUTION JACK LESTER KING UNIVERSITY OF CALIFORNIA, SANTA BARBARA This paper is dedicated to retiring University of California Professors Curt Stern and Everett R. Dempster. 1. Introduction Eleven decades of thought and work by Darwinian and neo-Darwinian scientists have produced a sophisticated and detailed structure of evolutionary ,theory and observations. In recent years, new techniques in molecular biology have led to new observations that appear to challenge some of the basic theorems of classical evolutionary theory, precipitating the current crisis in evolutionary thought. Building on morphological and paleontological observations, genetic experimentation, logical arguments, and upon mathematical models requiring simplifying assumptions, neo-Darwinian theorists have been able to make some remarkable predictions, some of which, unfortunately, have proven to be inaccurate. Well-known examples are the prediction that most genes in natural populations must be monomorphic [34], and the calculation that a species could evolve at a maximum rate of the order of one allele substitution per 300 genera- tions [13]. It is now known that a large proportion of gene loci are polymorphic in most species [28], and that evolutionary genetic substitutions occur in the human line, for instance, at a rate of about 50 nucleotide changes per generation [20], [24], [25], [26]. The puzzling observation [21], [40], [46], that homologous proteins in different species evolve at nearly constant rates is very difficult to account for with classical evolutionary theory, and at the very least gives a solid indication that there are qualitative differences between the ways molecules evolve and the ways morphological structures evolve. -

Microevolution and the Genetics of Populations Microevolution Refers to Varieties Within a Given Type
Chapter 8: Evolution Lesson 8.3: Microevolution and the Genetics of Populations Microevolution refers to varieties within a given type. Change happens within a group, but the descendant is clearly of the same type as the ancestor. This might better be called variation, or adaptation, but the changes are "horizontal" in effect, not "vertical." Such changes might be accomplished by "natural selection," in which a trait within the present variety is selected as the best for a given set of conditions, or accomplished by "artificial selection," such as when dog breeders produce a new breed of dog. Lesson Objectives ● Distinguish what is microevolution and how it affects changes in populations. ● Define gene pool, and explain how to calculate allele frequencies. ● State the Hardy-Weinberg theorem ● Identify the five forces of evolution. Vocabulary ● adaptive radiation ● gene pool ● migration ● allele frequency ● genetic drift ● mutation ● artificial selection ● Hardy-Weinberg theorem ● natural selection ● directional selection ● macroevolution ● population genetics ● disruptive selection ● microevolution ● stabilizing selection ● gene flow Introduction Darwin knew that heritable variations are needed for evolution to occur. However, he knew nothing about Mendel’s laws of genetics. Mendel’s laws were rediscovered in the early 1900s. Only then could scientists fully understand the process of evolution. Microevolution is how individual traits within a population change over time. In order for a population to change, some things must be assumed to be true. In other words, there must be some sort of process happening that causes microevolution. The five ways alleles within a population change over time are natural selection, migration (gene flow), mating, mutations, or genetic drift. -

Experiments on the Role of Deleterious Mutations As Stepping Stones In
Experiments on the role of deleterious mutations as PNAS PLUS stepping stones in adaptive evolution Arthur W. Covert IIIa,b,c,d, Richard E. Lenskia,c,e,1, Claus O. Wilkec,d, and Charles Ofriaa,b,c,1 aProgram in Ecology, Evolutionary Biology and Behavior, Departments of bComputer Science and Engineering and eMicrobiology and Molecular Genetics, and cBEACON Center for the Study of Evolution in Action, Michigan State University, East Lansing, MI 48824; and dCenter for Computational Biology, Institute for Cellular and Molecular Biology, and Department of Integrative Biology, University of Texas at Austin, Austin, TX 78712 Contributed by Richard E. Lenski, July 16, 2013 (sent for review April 22, 2013) Many evolutionary studies assume that deleterious mutations spread unless they are tightly linked (5, 6). In an asexual pop- necessarily impede adaptive evolution. However, a later mutation ulation, however, the fortuitous combination, once formed, will that is conditionally beneficial may interact with a deleterious be stably inherited, thereby providing a simple way to traverse a predecessor before it is eliminated, thereby providing access to fitness valley (5–10). adaptations that might otherwise be inaccessible. It is unknown Previous research on a variety of systems, both biological and whether such sign-epistatic recoveries are inconsequential events computational, has documented many examples of epistatic inter- or an important factor in evolution, owing to the difficulty of actions between mutations (3, 6, 11, 14–24), including a few cases monitoring the effects and fates of all mutations during experi- of mutational pairs that are individually deleterious but jointly ments with biological organisms. -

Patterns and Power of Phenotypic Selection in Nature
Articles Patterns and Power of Phenotypic Selection in Nature JOEL G. KINGSOLVER AND DAVID W. PFENNIG Phenotypic selection occurs when individuals with certain characteristics produce more surviving offspring than individuals with other characteristics. Although selection is regarded as the chief engine of evolutionary change, scientists have only recently begun to measure its action in the wild. These studies raise numerous questions: How strong is selection, and do different types of traits experience different patterns of selection? Is selection on traits that affect mating success as strong as selection on traits that affect survival? Does selection tend to favor larger body size, and, if so, what are its consequences? We explore these questions and discuss the pitfalls and future prospects of measuring selection in natural populations. Keywords: adaptive landscape, Cope’s rule, natural selection, rapid evolution, sexual selection henotypic selection occurs when individuals with selection on traits that affect survival stronger than on those Pdifferent characteristics (i.e., different phenotypes) that affect only mating success? In this article, we explore these differ in their survival, fecundity, or mating success. The idea and other questions about the patterns and power of phe- of phenotypic selection traces back to Darwin and Wallace notypic selection in nature. (1858), and selection is widely accepted as the primary cause of adaptive evolution within natural populations.Yet Darwin What is selection, and how does it work? never attempted to measure selection in nature, and in the Selection is the nonrandom differential survival or repro- century following the publication of On the Origin of Species duction of phenotypically different individuals. -

The Neutral Theory of Molecular Evolution
Copyright 2000 by the Genetics Society of America Perspectives Anecdotal, Historical and Critical Commentaries on Genetics Edited by James F. Crow and William F. Dove Thomas H. Jukes (1906±1999) James F. Crow Genetics Department, University of Wisconsin, Madison, Wisconsin 53706 OM Jukes accepted our invitation to write a Perspec- duced an immediate outcry from traditional students Ttives on the early history of molecular evolution, of evolution, undoubtedly abetted by the title. In the and in August 1999 he sent a rough beginning con- ensuing polemics, Kimura played the major role. King taining some now-forgotten early history. He planned an died prematurely in 1983 and Jukes wrote mainly about extensive revision and continuation, but on November 1 other things, although he did participate in one joint his death intervened. We have decided to publish his paper (Jukes and Kimura 1984). One of his interests early draft, realizing that it was but a start toward the was the evolution of the genetic code (Jukes 1983). I article that he had planned. particularly liked his showing how, in an orderly sequen- Tom, along with Jack L. King and Motoo Kimura, tial way, mutation pressure in the codon and anti-codon formulated the neutral theory of molecular evolution. could produce the unexpected codes in bacteria and Earlier, the idea had been foreshadowed by Sueoka mitochondria (Jukes 1985). He also developed a widely (1962) and Freese (1962). They had each suggested used correction for multiple undetected changes in evo- mutation pressure of near-neutral changes to account lutionary base substitutions (Jukes and Cantor 1969). -

Population Genetics of Neutral Mutations in Exponentially
The Annals of Applied Probability 2013, Vol. 23, No. 1, 230–250 DOI: 10.1214/11-AAP824 c Institute of Mathematical Statistics, 2013 POPULATION GENETICS OF NEUTRAL MUTATIONS IN EXPONENTIALLY GROWING CANCER CELL POPULATIONS By Rick Durrett1 Duke University In order to analyze data from cancer genome sequencing projects, we need to be able to distinguish causative, or “driver,” mutations from “passenger” mutations that have no selective effect. Toward this end, we prove results concerning the frequency of neutural mutations in exponentially growing multitype branching processes that have been widely used in cancer modeling. Our results yield a simple new population genetics result for the site frequency spectrum of a sample from an exponentially growing population. 1. Introduction. It is widely accepted that cancers result from an accu- mulation of mutations that increase the fitness of tumor cells compared to the cells that surround them. A number of studies [Sj¨oblom et al. (2006), Wood et al. (2007), Parsons et al. (2008), The Cancer Genome Atlas (2008) and Jones et al. (2008, 2010)] have sequenced the genomes of tumors in or- der to find the causative or “driver” mutations. However, due to the large number of genes being sequenced, one also finds a large number of “passen- ger” mutations that are genetically neutral and hence have no role in the disease. To explain the issues involved in distinguishing the two types of mutations, it is useful to take a look at a data set. Wood et al. (2007) did a “discovery” screen in which 18,191 genes were sequenced in 11 colorectal cancers, and then a “validation” screen in which the top candidates were sequenced in 96 additional tumors. -
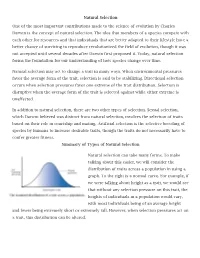
Natural Selection One of the Most Important Contributions Made to The
Natural Selection One of the most important contributions made to the science of evolution by Charles Darwin is the concept of natural selection. The idea that members of a species compete with each other for resources and that individuals that are better adapted to their lifestyle have a better chance of surviving to reproduce revolutionized the field of evolution, though it was not accepted until several decades after Darwin first proposed it. Today, natural selection forms the foundation for our understanding of how species change over time. Natural selection may act to change a trait in many ways. When environmental pressures favor the average form of the trait, selection is said to be stabilizing. Directional selection occurs when selection pressures favor one extreme of the trait distribution. Selection is disruptive when the average form of the trait is selected against while either extreme is unaffected. In addition to natural selection, there are two other types of selection. Sexual selection, which Darwin believed was distinct from natural selection, involves the selection of traits based on their role in courtship and mating. Artificial selection is the selective breeding of species by humans to increase desirable traits, though the traits do not necessarily have to confer greater fitness. Summary of Types of Natural Selection Natural selection can take many forms. To make talking about this easier, we will consider the distribution of traits across a population in using a graph. To the right is a normal curve. For example, if we were talking about height as a trait, we would see that without any selection pressure on this trait, the heights of individuals in a population would vary, with most individuals being of an average height and fewer being extremely short or extremely tall. -

Effectively Neutral Mutations
Proc. Natl. Acad. Sci. USA Vol. 76, No. 7, pp. 3440-3444, July 1979 Genetics Model of effectively neutral mutations in which selective constraint is incorporated (molecular evolution/protein polymorphism/population genetics/neutral mutation theory) MOTOO KIMURA National Institute of Genetics, Mishima 411, Japan Contributed by Motoo Kimura, April 25, 1979 ABSTRACT Based on the idea that selective neutrality is however, Ohta's model has a drawback in that it cannot ac- the limit when the selective disadvantage becomes indefinitely commodate enough mutations that behave effectively as neutral small, a model of neutral (and nearly neutral) mutations is pro- when the population size gets large. This difficulty can be posed that assumes that the selection coefficient (s') against the overcome by assuming that the selection coefficients follow a mutant at various sites within a cistron (gene) follows a r dis- r distribution. tribution; Ass') = aPe-as's.-l/P(,B), in which a = fi/i and s' is the mean selection coefficient against the mutants (?'> 0; 1 _ ft > 0). The mutation rate for alleles whose selection coef- MODEL OF EFFECTIVELY NEUTRAL ficients s' lie in the range between 0 and 1/(2Ne), in which Ne MUTATIONS is the effective population size, is termed the effectively neutral mutation rate (denoted by ye). Using the model of "infinite Let us assume that the frequency distribution of the selective sites" in population genetics, formulas are derived giving the disadvantage (denoted by s') of mutants among different sites average heterozygosity (he) and evolutionary rate per generation follows the r distribution (kg) in terms of mutant substitutions. -

Supplementary Information for Vyssotski, 2013
Supplementary Information for Vyssotski, 2013 www.evolocus.com/evolocus/v1/evolocus-01-013-s.pdf Evolocus Supplementary Information for Transgenerational epigenetic compensation and sex u al d imorph ism 1,2,3 Dmitri L. Vyssotski Published 17 May 2013, Evolocus 1, 13-18 (2013): http://www.evolocus.com/evolocus/v1/evolocus-01-013.pdf In natu ral popu lation transgenerational epigenetic compensation is by unknown mechanisms, the mechanisms those were difficult to generated in h omoz y gou s mu tants, mainly in males. It d oes not imagine, and this possibility was abandoned for over 47 years. immed iately affect th e ph enoty pe of h omoz y gou s mu tants, w h ere The higher variability in males can be a result of a not so good it w as generated . Transgenerational epigenetic compensation is canalization of their ontogenesis, and the canalization of localiz ed not in th e same locu s as original mu tation and it is ontogenesis is modulated by sex hormones. Thus, we can d ominant, b ecau se it affects th e ph enoty pe ev en if it is receiv ed imagine that the canalization of ontogenesis in males is from one parent. In fu rth er generations, starting from F 2, it is more artificially decreased in comparison with females by means of ex pressed in females th an in males: it increases fitness of hormonal differences. h omozy gou s mu tant females, d ecreases fitness of w ild ty pe The shift of the female distribution to the right was not females and h as w eak effect on h eterozy gou s females. -

Stabilizing Selection Shapes Variation in Phenotypic Plasticity
bioRxiv preprint doi: https://doi.org/10.1101/2021.07.29.454146; this version posted July 29, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 Stabilizing selection shapes variation in phenotypic plasticity 2 3 4 5 Dörthe Becker1,2,*, 6 Karen Barnard-Kubow1,3, 7 Robert Porter1, 8 Austin Edwards1,4, 9 Erin Voss1,5, 10 Andrew P. Beckerman2, 11 Alan O. Bergland1,* 12 13 14 1Department of Biology; University of Virginia, Charlottesville/VA, USA 15 2Department of Animal and Plant Sciences; University of Sheffield, Sheffield, UK 16 3Department of Biology, James Madison University, Harrisonburg/VA, USA 17 4Biological Imaging Development CoLab, University of California San Francisco, 18 San Francisco/CA, USA 19 5Department of Integrative Biology, University of California Berkeley, 20 Berkeley/CA 21 22 * to whom correspondence should be addressed: 23 DB ([email protected]), AOB ([email protected]) 24 25 26 27 Keywords: stabilizing selection; phenotypic plasticity; heritability; Daphnia; 28 defense morphologies 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.07.29.454146; this version posted July 29, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 29 Abstract 30 The adaptive nature of phenotypic plasticity is widely documented in natural 31 populations. However, little is known about the evolutionary forces that shape 32 genetic variation in plasticity within populations. Here we empirically address this 33 issue by testing the hypothesis that stabilizing selection shapes genetic variation 34 in the anti-predator developmental plasticity of Daphnia pulex. -

Selection on Breeding Date and Body Size in Colonizing Coho Salmon, Oncorhynchus Kisutch
Molecular Ecology (2010) 19, 2562–2573 doi: 10.1111/j.1365-294X.2010.04652.x Selection on breeding date and body size in colonizing coho salmon, Oncorhynchus kisutch J. H. ANDERSON,* P. L. FAULDS,† W. I. ATLAS,‡ G. R. PESS§ and T. P. QUINN* *School of Aquatic and Fishery Sciences, University of Washington, Box 355020, Seattle, WA 98195, USA, †Seattle Public Utilities, 700 5th Avenue, Suite 4900, Seattle, WA, 98124, USA, ‡Department of Biological Sciences, Simon Fraser University, 8888 University Drive, Burnaby, BC, Canada V5A 1S6, §National Marine Fisheries Service, Northwest Fisheries Science Center, 2725 Montlake Boulevard East, Seattle, WA 98195, USA Abstract Selection during the colonization of new habitat is critical to the process of local adaptation, but has rarely been studied. We measured the form, direction, and strength of selection on body size and date of arrival to the breeding grounds over the first three cohorts (2003–2005) of a coho salmon (Oncorhynchus kisutch) population colonizing 33 km of habitat made accessible by modification of Landsburg Diversion Dam, on the Cedar River, Washington, USA. Salmon were sampled as they bypassed the dam, parentage was assigned based on genotypes from 10 microsatellite loci, and standardized selection gradients were calculated using the number of returning adult offspring as the fitness metric. Larger fish in both sexes produced more adult offspring, and the magnitude of the effect increased in subsequent years for males, suggesting that low densities attenuated traditional size-biased intrasexual competition. For both sexes, directional selection favoured early breeders in 2003, but stabilizing selection on breeding date was observed in 2004 and 2005.