Multiple-Scale Habitat Relationships of Tidal-Marsh Breeding Birds in the San Francisco Bay Estuary
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distribution and Abundance
DISTRIBUTION AND ABUNDANCE IN RELATION TO HABITAT AND LANDSCAPE FEATURES AND NEST SITE CHARACTERISTICS OF CALIFORNIA BLACK RAIL (Laterallus jamaicensis coturniculus) IN THE SAN FRANCISCO BAY ESTUARY FINAL REPORT To the U.S. Fish & Wildlife Service March 2002 Hildie Spautz* and Nadav Nur, PhD Point Reyes Bird Observatory 4990 Shoreline Highway Stinson Beach, CA 94970 *corresponding author contact: [email protected] PRBO Black Rail Report to FWS 2 EXECUTIVE SUMMARY We conducted surveys for California Black Rails (Laterallus jamaicensis coturniculus) at 34 tidal salt marshes in San Pablo Bay, Suisun Bay, northern San Francisco Bay and western Marin County in 2000 and 2001 with the aims of: 1) providing the best current information on distribution and abundance of Black Rails, marsh by marsh, and total population size per bay region, 2) identifying vegetation, habitat, and landscape features that predict the presence of black rails, and 3) summarizing information on nesting and nest site characteristics. Abundance indices were higher at 8 marshes than in 1996 and earlier surveys, and lower in 4 others; with two showing no overall change. Of 13 marshes surveyed for the first time, Black Rails were detected at 7 sites. The absolute density calculated using the program DISTANCE averaged 2.63 (± 1.05 [S.E.]) birds/ha in San Pablo Bay and 3.44 birds/ha (± 0.73) in Suisun Bay. At each survey point we collected information on vegetation cover and structure, and calculated landscape metrics using ArcView GIS. We analyzed Black Rail presence or absence by first analyzing differences among marshes, and then by analyzing factors that influence detection of rails at each survey station. -

2010-2011 California Regulations for Waterfowl and Upland Game Hunting, Public Lands
Table of Contents CALIFORNIA General Information Contacting DFG ....................................... 2 10-11 Licenses, Stamps, & Permits................... 3 Waterfowl & Upland Shoot Time Tables ................................... 4 Game Hunting and Unlawful Activities .......................... 6 Hunting & Other Public Uses on State & Federal Waterfowl Hunting Lands Regulations Summary of Changes for 10-11 ............... 7 Seasons and Limits ................................. 9 Effective July 1, 2010 - June 30, 2011 Waterfowl Consumption Health except as noted. Warnings ............................................. 12 State of California Special Goose Hunt Area Maps ............ 14 Governor Arnold Schwarzenegger Waterfowl Zone Map ....Inside Back Cover Natural Resources Agency Upland Game Bird, Small Game Secretary Lester A. Snow Mammal, and Crow Hunting Regulation Summary ............................. 16 Fish and Game Commission President Jim Kellogg Seasons & Limits Table ......................... 17 Vice President Richard B. Rogers Hunt Zones ............................................ 18 Commissioner Michael Sutton Hunting and Other Public Uses on Commissioner Daniel W. Richards State and Federal Areas Acting Executive Director Jon Fischer Reservation System .............................. 20 General Public Use Activities on Department of Fish and Game Director John McCamman All State Wildlife Areas ....................... 23 Hunting, Firearms, and Archery Alternate communication formats are available upon request. If reasonable -

550. Regulations for General Public Use Activities on All State Wildlife Areas Listed
550. Regulations for General Public Use Activities on All State Wildlife Areas Listed Below. (a) State Wildlife Areas: (1) Antelope Valley Wildlife Area (Sierra County) (Type C); (2) Ash Creek Wildlife Area (Lassen and Modoc counties) (Type B); (3) Bass Hill Wildlife Area (Lassen County), including the Egan Management Unit (Type C); (4) Battle Creek Wildlife Area (Shasta and Tehama counties); (5) Big Lagoon Wildlife Area (Humboldt County) (Type C); (6) Big Sandy Wildlife Area (Monterey and San Luis Obispo counties) (Type C); (7) Biscar Wildlife Area (Lassen County) (Type C); (8) Buttermilk Country Wildlife Area (Inyo County) (Type C); (9) Butte Valley Wildlife Area (Siskiyou County) (Type B); (10) Cache Creek Wildlife Area (Colusa and Lake counties), including the Destanella Flat and Harley Gulch management units (Type C); (11) Camp Cady Wildlife Area (San Bernadino County) (Type C); (12) Cantara/Ney Springs Wildlife Area (Siskiyou County) (Type C); (13) Cedar Roughs Wildlife Area (Napa County) (Type C); (14) Cinder Flats Wildlife Area (Shasta County) (Type C); (15) Collins Eddy Wildlife Area (Sutter and Yolo counties) (Type C); (16) Colusa Bypass Wildlife Area (Colusa County) (Type C); (17) Coon Hollow Wildlife Area (Butte County) (Type C); (18) Cottonwood Creek Wildlife Area (Merced County), including the Upper Cottonwood and Lower Cottonwood management units (Type C); (19) Crescent City Marsh Wildlife Area (Del Norte County); (20) Crocker Meadow Wildlife Area (Plumas County) (Type C); (21) Daugherty Hill Wildlife Area (Yuba County) -

Bel Marin Keys
Prepared by: SR-37 Corridor Adaptation Study Prepared for: SEGMENT A-1 ADAPTATION STRATEGIES This corridor adaptation study is funded through Senate Bill 1 (SB 1) 2018 Adaptation Planning Grant Table of Contents Acknowledgments 5 Glossary of Terms 7 Figure 1.A Key Issues Along SR-37 in Marin 9 1.01 Introduction 10 1.02 Grant Purpose 11 1.03 Study Outreach 11 1.04 Basis of Study 12 1.05 Segment A-1 Challenges 12 Figure 1.B Future Sea Level Rise Inundation 13 Figure 1.C Understanding Roadway Elevation Requirements 14 1.06 Segment A-1 Opportunities 15 1.07 Segment A-1 Related Studies Key Findings 16 Figure 1.D Lower Novato Creek Watershed Flow 18 Figure 1.E SR-37/US-101 Interchange Considerations 19 Figure 1.F Corridor Land Use 21 1.08 Visions for Resiliency 26 Figure 1.I Vision 1 “Hold the Line” 28 Figure 1.J Vision 2 Proposed Section 29 Figure 1.K Vision 2 “Move the Line” 30 Figure 1.L Vision 3 “Multi-Bene icial” 32 Figure 1.M Vision 4 (Proposed) Section 33 Figure 1.N Vision 4 “Buying Time” 34 1.09 Next Steps 35 1.10 Project Stakeholders 37 Figure 1.O Long Term Restoration Novato Creek 38 Figure 1.P Maximize Habitat, Minimize Risk Strategy 39 Figure 1.Q Existing Public Land Ownership 40 Figure 1.R Vulnerability Map of Novato 41 2. Summary of Studies 46 Transportation Studies 46 2.01 Highway 37 Stewardship Study (Case Study) 46 2.02 The State Route 37 Transportation Concept Report 47 2.03 SR-37 Transportation and Sea Level Rise Corridor Plan 48 2.04 SR-37 Segment A Sea Level Rise Corridor Improvement Study H 50 2.05 The Project Study Report -

Fishes-Of-The-Salish-Sea-Pp18.Pdf
NOAA Professional Paper NMFS 18 Fishes of the Salish Sea: a compilation and distributional analysis Theodore W. Pietsch James W. Orr September 2015 U.S. Department of Commerce NOAA Professional Penny Pritzker Secretary of Commerce Papers NMFS National Oceanic and Atmospheric Administration Kathryn D. Sullivan Scientifi c Editor Administrator Richard Langton National Marine Fisheries Service National Marine Northeast Fisheries Science Center Fisheries Service Maine Field Station Eileen Sobeck 17 Godfrey Drive, Suite 1 Assistant Administrator Orono, Maine 04473 for Fisheries Associate Editor Kathryn Dennis National Marine Fisheries Service Offi ce of Science and Technology Fisheries Research and Monitoring Division 1845 Wasp Blvd., Bldg. 178 Honolulu, Hawaii 96818 Managing Editor Shelley Arenas National Marine Fisheries Service Scientifi c Publications Offi ce 7600 Sand Point Way NE Seattle, Washington 98115 Editorial Committee Ann C. Matarese National Marine Fisheries Service James W. Orr National Marine Fisheries Service - The NOAA Professional Paper NMFS (ISSN 1931-4590) series is published by the Scientifi c Publications Offi ce, National Marine Fisheries Service, The NOAA Professional Paper NMFS series carries peer-reviewed, lengthy original NOAA, 7600 Sand Point Way NE, research reports, taxonomic keys, species synopses, fl ora and fauna studies, and data- Seattle, WA 98115. intensive reports on investigations in fi shery science, engineering, and economics. The Secretary of Commerce has Copies of the NOAA Professional Paper NMFS series are available free in limited determined that the publication of numbers to government agencies, both federal and state. They are also available in this series is necessary in the transac- exchange for other scientifi c and technical publications in the marine sciences. -

The Draft Black Point Community Plan Was Established As a Result of the Process to Update the 1978 Black Point Community Plan. O
April 20, 2016 Draft Black Point Community Plan Summary of Key Changes The Draft Black Point Community Plan was established as a result of the process to update the 1978 Black Point Community Plan. On March 8, 2016, the Marin County Board of Supervisors directed staff to develop a separate community plan for the Black Point community. The updated Plan is based on the Planning Commission recommended draft of the Black Point/Green Point Communities Plan, which has been edited to remove content and policies not relevant to Black Point. A summary of the key changes are as follows: • Renamed the Draft Black Point/Green Point Community Plan to the Draft Black Point Community Plan; • Updated content to refer to Black Point throughout document; • Created a separate community plan boundary for Black Point; • Noted that all census demographic information for the community is based on the Black Point – Green Point Census Designated Place, which includes Green Point. Specific information for Black Point is not available at this time; • Proposed policies regulating home size and setbacks have been removed at the direction of the Board of Supervisors; and • Updated land use and zoning information in the Community Character and Land Use chapter. A separate version of the Draft is also available showing all changes in strike-out and underline format on www.marincounty.org/blackpoint or http://www.marincounty.org/~/media/files/departments/cd/planning/community- planning/blackpoint/blackpoint_draft_tracked_04152016.pdf?la=en. Black Point Community Plan -

California Clapper Rail (Rallus Longirostris Obsoletus) Population Monitoring: 2005-2008
California Clapper Rail (Rallus longirostris obsoletus) Population monitoring: 2005-2008 Leonard Liu, Julian Wood, Nadav Nur, Diana Stralberg, and Mark Herzog PRBO Conservation Science 3820 Cypress Drive #11, Petaluma, CA 94954 FINAL September 29, 2009 Prepared for: California Department of Fish and Game 4001 N. Wilson Way Stockton, CA 95205 PRBO California Clapper Rail Monitoring Report, 2005-2008 FINAL Abstract PRBO Conservation Science conducted call-count surveys for California Clapper Rail ( Rallus longirostrus obsoletus ) at 53 sites throughout the San Francisco Bay Estuary from 2005 through 2008. To maximize the spatial coverage of sites, surveys were coordinated with partners conducting call-count surveys (Avocet Research Associates, California Department of Fish and Game, California Coastal Conservancy’s Invasive Spartina Project, and U.S. Fish and Wildlife Service) resulting in a total of 180 sites surveyed, covering all high-quality habitat and the majority of lower-quality habitat . We estimated annual site-specific density estimates using distance sampling and program DISTANCE and detected a negative short-term trend of -20.6% (±3.8%) from 2005 through 2008. There were no significant changes in densities from 2005 to 2006 or from 2006 to 2007. From 2007 to 2008, an Estuary-wide negative change was detected (-46.0%, ±6.75%) which was driven by a dramatic decrease in South San Francisco Bay (-57.4% ±5.0%). We tested the power to detect a 10-year trend for five different monitoring scenarios and found that detecting an Estuary-wide negative trend of 13.9% or greater over a 10-year period with 80% power is possible under the current monitoring design (90 sites/year with effort allocated equally among sites). -

Department of Fish and Game Marine Resources Operations
Report for the month of May 1966 Item Type monograph Publisher California Department of Fish and Game, Marine Resources Operations Download date 03/10/2021 16:35:02 Link to Item http://hdl.handle.net/1834/18921 THE RESOURCES AGENCY OF CALIFORNIA DEPARTMENT OF FISH AND GAME MARINE RESOURCES OPERATIONS REPORT FOR THE MONTH OF MAY 1966 May jack mackerel landings (4,700 tons) were the largest in the history of the fishery for thi~ month. Landings for the year this far are still 3,500 tons behind the 10-year average. The Fisheries Resources 'Sea Survey' Project (Bartlett M63R) was approved effective March 8, 1966. First personnel were assigned effective· April 1, 1966. Over 3,150 elephant 'seals were counted on two Southern California islands during May: 3,090 on San Miguel Island, May 3, and 80 on Santa Barbara Island, May 24. A ,tagged California halibut was recovered 258 days and 115" miles from its release date and locality. Very few of our tagged halibut have shown move ments of such magnitude in the past. - 2 - Pelagic Fish .................... ~ ~ , 3 Tuna 4 Sportfish II ••••••••••••••••••• ! • eo ••• 5 Special Prbjects '......................... 7 Biostatistics 8 Miscellaneous 9 - 3 - 1. PELAGIC FISH A. Fishery May January 1 -May 31 10--yr. --mean Landings in tons 1966* 1965 1966* 1965 -1955..:.1964 Species Anchovy 16 174 17,094 588 5,269 Mackerel, jack 4,700 1,979 8,615 5,843 12,153 Mackerel, Pacific 150 21 524 139 5,238 Sardines 30 54 103 393 1,523 Squid 1 352 2,312 2,997 -2;624 4,897 2,580 28,648 9,960 26,807 *Estimated. -

California Clapper Rail (Rallus Longirostris Obsoletus) Population Monitoring: 2005-2011 Final Technical Report
California Clapper Rail (Rallus longirostris obsoletus) Population monitoring: 2005-2011 Final Technical Report Leonard Liu, Julian Wood, Nadav Nur, Leo Salas, and Dennis Jongsomjit PRBO Conservation Science 3820 Cypress Drive #11, Petaluma, CA 94954 California Clapper Rail Population Monitoring 2005-2011 Final Report Table of Contents ACKNOWLEDGMENTS ....................................................................................... 3 EXECUTIVE SUMMARY ....................................................................................... 4 INTRODUCTION .................................................................................................. 6 METHODS .............................................................................................................. 9 FIELD SURVEYS .................................................................................................................... 9 ANALYSES ......................................................................................................................... 10 MODEL APPROACH ............................................................................................................. 11 ECOLOGICAL MODEL ........................................................................................................... 11 Detection Sub-model. ............................................................................................................................. 11 Abundance Sub-model. ......................................................................................................................... -
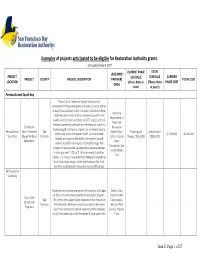
Updated Draft Project List And
Examples of projects anticipated to be eligible for Restoration Authority grants. Last updated June 9, 2017 TOTAL LEAD (AND CURRENT PHASE PROJECT SCHEDULE CURRENT PROJECT COUNTY PROJECT DESCRIPTION PARTNER) SCHEDULE TOTAL COST LOCATION (Phase; dates or (Phase; dates PHASE COST ORGS years) or years) Peninsula and South Bay Phase 2 of the Yosemite Slough Restoration & Development Project will green and open a 21‐acre section of waterfront parkland in San Francisco's Candlestick Point California State Recreation Area that has remained closed to the Department of public since the park's inception in 1977, add 21 acres of Parks and restored waterfront parklands and recreational space in a Candlestick Recreation disadvantaged community, improve air and water quality, Peninsula and Point ‐ Yosemite San (State Parks), Planning and Construction: reduce and clean stormwater runoff, provide wildlife $1,300,000 $6,400,000 South Bay Slough Wetland Francisco California State Design: 2016‐2018 2018‐2019 habitat, and improve the ability of the park's natural Restoration Parks systems to buffer the impacts of climate change. The Foundation; San project will also provide valuable public access amenities Francisco Bay including a new 1,100 sq. ft. zero net energy Education Trail Center, 1.1 miles of new waterfront biking and pedestrian trails (including a section of the San Francisco Bay Trail), and ADA‐accessible park viewing and picnic/BBQ areas. Peninsula and South Bay Environmental education programs for students of all ages Golden Gate at the Crissy Field Center related to restoration projects. National Parks Crissy Field San The Center offers place‐based exploration that focuses on Conservancy, Educational ‐ ‐ ‐ ‐ Francisco the interaction between humans and nature and makes National Park Programs use of the natural and cultural resources of the restored Service, Presidio Crissy Field wetland and the Tennessee Hollow watershed. -

2020 Ridgway's Rail Survey Report
California Ridgway’s Rail Surveys for the San Francisco Estuary Invasive Spartina Project 2020 Report to: The State Coastal Conservancy San Francisco Estuary Invasive Spartina Project 1515 Clay St., 10th Floor Oakland, CA 94612 Prepared by: Olofson Environmental, Inc. 1001 42nd Street, Suite 230 Oakland, California 94608 Contact: [email protected] February 1, 2021 ACKNOWLEDGEMENTS This report was designed and prepared under the direction of Jen McBroom, the Invasive Spartina Project Ridgway’s Rail Monitoring Manager, with considerable hard work by other OEI biologists and staff, including Brian Ort, Jeanne Hammond, Kevin Eng, Nate Deakers, Pim Laulikitnont, Simon Gunner, Stephanie Chen, Tobias Rohmer, Melanie Anderson, and Lindsay Faye. This report was prepared for the California Coastal Conservancy’s San Francisco Estuary Invasive Spartina Project Table of Contents 1. Introduction ...................................................................................................................................... 1 2. Study Area ......................................................................................................................................... 3 3. Methods ............................................................................................................................................. 5 3.1 Field Methods ........................................................................................................................ 5 3.2 Data Management ................................................................................................................ -

Land and Water Conservation Fund Program Funded Projects
Land and Water Conservation Fund Program Funded Projects Alameda County Alameda Creek Regional Park Alameda Creek-Coyote Hills Acquisition 1966/67 $819,997 Acquisition of 446 acres. Alameda Shoreline Park Alameda Shoreline Park 1978/79 $335,280 Picnic and play areas, trails. Alvarado Community Center Alvarado/Chavez Park Development 2001/02 $71,466 Development of a gazebo at Alvarado/Chavez Park to include renovated structure and new paint. Aquatic Park Aquatic Park Acquisition 1970/71 $24,633 Acquisition of 1.6 acre addition. Aquatic Park Development 1970/71 $154,581 Picnic areas, tot lot, roads, parking, irrigation, landscaping, and play fields. Aquatic Park Development 1976/77 $107,766 Play area, day camp, picnic area, landscaping, pathways. Ardenwood Park Ardenwood Regional Park Development 1983/84 $201,600 Development of access roads, parking, landscaping, restrooms, utilities and stabilization of farm buildings. Arroyo Swim Center Arroyo/Castro Valley Swim Development 1979/80 $81,280 Renovated swim pools at Castro Valley and Arroyo Swim Centers. Bateman Mall Park Bateman Park Development 1983/84 $37,800 Tot lot, lighting, irrigation, landscaping, paths. Big Break Regional Shoreline Vintage Parkway Creek Development 2003/04 $202,700 Development of a trail bridge across the Vintage Parkway Creek Channel. Caldecott Park Caldecott Park 1980/81 $202,400 Picnic areas, sports & play fields, trails, parking and restrooms. Caldecott Park Acquisition 1976/77 $151,756 57.1 acres acquired adjacent to the road for regional sports facility. Castro Valley Swim Center Arroyo/Castro Valley Swim Development 1979/80 $81,280 Renovated swim pools at Castro Valley and Arroyo Swim Centers. Centennial Park Centennial Park 1979/80 $165,408 Picnic areas, sports fields, play fields.