Free Executive Summary
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Geo-Heat Center Quarterly Bulletin Vol. 16, No. 4
Vol. 16, No. 4 OCTOBER 1995 GEO-HEAT CENTER QUARTERLY BULLETIN ISSN 0276-1084 A Quarterly Progress and Development Report on the Direct Utilization of Geothermal Resources CONTENTS Page PUBLISHED BY 20TH ANNIVERSARY OF THE GEO- 1 GEO-HEAT CENTER HEAT CENTER Oregon Institute of Technology P. J. Lienau and J. Lund 3201 Campus Drive Geo-Heat Center Klamath Falls, OR 97601 Phone: 541-885-1750 HISTORICAL IMPACTS OF 7 Email: [email protected] GEOTHERMAL RESOURCES ON THE PEOPLE OF All articles for the Bulletin are solicited. If you NORTH AMERICA wish to contribute a paper, please contact the J. Lund editor at the above address. Geo-Heat Center EDITOR COLLOCATED RESOURCES 15 T. Boyd Geo-Heat Center Paul J. Lienau and John W. Lund Typesetting and Layout – Donna Gibson KLAMATH FALLS SNOW 23 MELT SYSTEM B. Brown Consultant FUNDING PRAWN PARK – 27 The Bulletin is provided compliments of the Geo- TAUPO, NEW ZEALAND Heat Center. This material was prepared with the J. Lund and R. Klein support of the U.S. Department of Energy (DOE Geo-Heat Center Grant No. DE-FG07-90ID13040). However, any opinions, findings, conclusions, or GEOTHERMAL PIPELINE 30 recommendations expressed herein are those of Progress and Development Update the authors(s) and do not necessarily reflect the From the Geothermal Progress view of USDOE. Monitor SUBSCRIPTIONS Cover: (1) 1974 International Conference on The Bulletin is mailed free of charge. Please send Geothermal energy for Industrial, Agricultural and your name and address to the Geo-Heat Center for commercial-Residential Uses, held at OIT; (2) OIT experimental greenhouse; (3) aerial view of OIT addition to the mailing list. -

Lower and Middle Palaeolithic Leicestershire and Rutland: Progress and Potential by Anne Graf
Leic.Arch. Sept 2002 11/10/02 7:54 AM Page 1 Lower and Middle Palaeolithic Leicestershire and Rutland: progress and potential by Anne Graf Drawing on archaeological and geological research, recent finds, museum collections and borehole evidence, this paper describes the changing local landscape and environment round the Cromerian-period Bytham river valley, possibly the major early entry route to Britain from Europe; the succeeding pro-glacial Lake Harrison and present-day post-glacial rivers; and highlights deposits with Palaeolithic archaeological potential. Palaeolithic tools of quartzite, comparable with other Midlands raw materials, and of andesitic and Charnian tuff may suggest occupation from around 600,000 BC, while flint tools suggest at least intermittent post-Anglian occupation, followed by probable abandonment from after 160,000 BC until possible Devensian re-occupation by Neanderthalers post- 60,000 BC. Introduction The last twenty years have seen very exciting results for the Palaeolithic period in Leicestershire and Rutland. This paper discusses these results, which are placed in the contexts both of the local landscape changes, and of the growing national understanding of key Palaeolithic issues. The paper aims to provide a summary of current knowledge of the Lower and Middle Palaeolithic occupants and their environments and landscapes in these two counties, and to highlight potential for further investigation. The local, and many national, sites, to which references can be found in the bibliography, are presented in the timechart in illus. 1, with their relative positions within current archaeological and geological divisions and phases. Throughout the timechart and the rest of this paper ‘thousand [years] ago’ will be abbreviated to ‘ka’, while the oxygen isotope stages now used for international date correlations (see below) will also be abbreviated, so that, for example, OIS-16 indicates oxygen isotope stage 16. -

Drainage Network Reorganization Affecting the Nene and Welland Catchments of Eastern England As a Result of a Late Middle Pleistocene Glacial Advance
Received: 25 September 2017 | Revised: 9 May 2018 | Accepted: 14 May 2018 DOI: 10.1002/dep2.44 ORIGINAL RESEARCH ARTICLE Drainage network reorganization affecting the Nene and Welland catchments of eastern England as a result of a late Middle Pleistocene glacial advance Harry E. Langford Department of Geography, Birkbeck University of London, London, UK Abstract Britain’s latitude is critical for determining the southerly extent of western Euro- Correspondence pean ice sheet expansion. Palaeoclimate and palaeosea-level reconstruction in Bri- Harry E. Langford, Department of Geography, Birkbeck University of tain, however, is complicated by spatially discontinuous glacigenic deposits and London, Malet Street, London WC1E genetic and stratigraphic interpretations have relied on their lithological character- 7HX, UK. istics. This study adopted a sedimentary facies approach combined with geomor- Email: [email protected] phological analyses for investigating upper Middle Pleistocene deposits in two adjacent catchments: Nene and Welland. Detailed sedimentology determined not only genesis of ice-contact (Welland) and glaciolacustrine (Nene) deposits but also provided insight on lake surface water levels. The ice-contact deposits recorded a constant lake level at the same height as alluvial remnants upstream in the Wel- land valley. These alluvial remnants are located where present tributaries join the Welland, indicating they were formed where influent streams entered a former lake and therefore can be interpreted as fluvial terraces resulting from local base- level rise. The glaciolacustrine deposits in the Nene valley recorded fluctuating lake levels, indicating that altitudinally separated sand and gravel bodies coinci- dent with these fluctuations and located where influent streams entered the lake also can be interpreted as fluvial terraces, but resulting from fluctuating baselevel. -

Lund HISTORICAL IMPACTS of GEOTHERMAL RESOURCES ON
Lund HISTORICAL IMPACTS O F GEOTHERMAL RESOURCES ON THE PEOPLE O F NORTH AMERICA John W. Lund Geo-Heat Center, Oregon Institute of Technology Klamath Falls. Oregon, USA Key words: history, Indians, hot springs, volcanoes, legends ABSTRACT The Indians of North America considered hot springs as a sacred place where the "Great Spirit" lived, and thus were great believers in the miraculous healing powers of the heat and mineral waters. These areas were also known as neutral ground; where warriors could travel to and rest unmolested by other tribes. Even though archeological finds date Native American presence at hot springs for over 10,000 years, there is no recorded history prior to the arrival of the Europeans in the Many legends concerning geothermal activities are part of the Native American history, such as about Madame Pele, the Hawaiian goddess of volcanic fire, and the story of the battle between Skell and Llao describing the eruptions of Mt. Mazama (Crater Lake) and Shasta. Obsidian was one of the prized volcanic trading items used by the Indians for tools and weapons. INTRODUCTION \ The history of geothermal use prior to the industrial revolution, centers itself mainly in the volcanic region western I North America. A few isolated locations are also found in the middle and eastern pan of the continent. The use of the geothermal resources during this period was by Paleo-Indians and I Indians, the latter also referred as Native Americans. Even though archeological finds date Native American presence at hot springs for over years, there is no recorded history prior to the arrival of the Europeans in the Rock paintings (pictographs) and rack carvings (petroglyphs) are their only source Figure 1. -

Prospecting Glacial Ages and Paleoclimatic Reconstructions Northeastward of Nevado Coropuna (16◦ S, 73◦ W, 6377 M), Arid Tropical Andes
geosciences Article Prospecting Glacial Ages and Paleoclimatic Reconstructions Northeastward of Nevado Coropuna (16◦ S, 73◦ W, 6377 m), Arid Tropical Andes Jose Úbeda 1,2,3,* ID , Martí Bonshoms 2, Joshua Iparraguirre 1, Lucía Sáez 3, Ramón de la Fuente 3, Lila Janssen 3, Ronald Concha 1, Pool Vásquez 1 and Pablo Masías 1 1 Instituto Geológico Minero y Metalúrgico, Av. Canadá 1470, San Borja 15034, Peru; [email protected] (J.I.); [email protected] (R.C.); [email protected] (P.V.); [email protected] (P.M.) 2 Grupo de Investigación en Geografía Física de Alta Montaña, Departamento de Geografía, Universidad Complutense de Madrid, 28040 Madrid, Spain; [email protected] 3 Guías de Espeleología y Montaña (Speleology and Mountain Guides), Casilla del Mortero, Torremocha de Jarama, 28189 Madrid, Spain; [email protected] (L.S.); [email protected] (R.d.l.F.); [email protected] (L.J.) * Correspondence: [email protected]; Tel.: +34-656408790 Received: 19 July 2018; Accepted: 15 August 2018; Published: 20 August 2018 Abstract: This work investigates the timing, paleoclimatic framework and inter-hemispheric teleconnections inferred from the glaciers last maximum extension and the deglaciation onset in the Arid Tropical Andes. A study area was selected to the northeastward of the Nevado Coropuna, the volcano currently covered by the largest tropical glacier on Earth. The current glacier extent, the moraines deposited in the past and paleoglaciers at their maximum extension have been mapped. The present and past Equilibrium Line Altitudes (ELA and paleoELA) have been reconstructed and the chlorine-36 ages have been calculated, for preliminary absolute dating of glacial and volcanic processes. -

Early Middle Pleistocene Drainage in Southern Central England
Netherlands Journal of Geosciences —– Geologie en Mijnbouw | 93 – 4 | 135-145 | 2014 doi: 10.1017/njg.2014.25 Early Middle Pleistocene drainage in southern central England R.K. Belshaw1, P.L. Gibbard2,*,J.B.Murton3 &D.K.Murton2 1 6a Ipswich Road, Norwich NR2 2LP, United Kingdom 2 Cambridge Quaternary, Department of Geography, University of Cambridge, Cambridge CB2 3EN, United Kingdom 3 Permafrost Laboratory, Department of Geography, University of Sussex, Brighton BN1 9QJ, United Kingdom * Corresponding author. Email: [email protected] Manuscript received: 8 April 2014, accepted: 30 June 2014 Abstract The fluvial sequences of the Milton and the Letchworth formations in the south Midlands of England and neighbouring regions represent at least two pre-existing rivers, the Milton and Brigstock streams, underlying Middle Pleistocene glacial sediments. The Milton Formation includes sand sourced from the Midlands bedrock. This implies that both streams were aligned in a northwest to southeast direction. This direction parallels the contem- poraneous courses of the rivers Thames and Trent, the former turning towards the east and northeast to enter the North Sea. Their alignments indicate that the Milton and Letchworth streams formed left-bank tributaries of the Thames, joining the river in Hertfordshire and Essex, as illus- trated in the article. This reconstruction has important implications for the interpretation of the proto-Soar river of the south Midlands, represented by the Baginton Formation. Although originally thought to represent a late Middle Pleistocene line, this southwest to northeast aligned system was reinterpreted as the headwaters of a pre-Anglian ‘Bytham river’, a1ligned towards East Anglia. However, recent work has shown that this river could not have existed in the pre-Anglian since there is no link between the Midlands and East Anglian spreads. -

Baker-Fritzqsr2015.Pdf
Quaternary Science Reviews 124 (2015) 31e47 Contents lists available at ScienceDirect Quaternary Science Reviews journal homepage: www.elsevier.com/locate/quascirev Invited review Nature and causes of Quaternary climate variation of tropical South America Paul A. Baker a, b, *, Sherilyn C. Fritz c, d a Division of Earth and Ocean Sciences, Duke University, Durham, NC 27708, USA b Yachay Tech University, School of Geological Sciences and Engineering, San Miguel de Urcuqui, Hacienda San Jose, Imbabura, Ecuador c Department of Earth and Atmospheric Sciences, University of Nebraska e Lincoln, Lincoln, NE 68588-0340, USA d School of Biological Sciences, University of Nebraska e Lincoln, Lincoln, NE 68588-0340, USA article info abstract Article history: This selective review of the Quaternary paleoclimate of the South American summer monsoon (SASM) Received 11 February 2015 domain presents viewpoints regarding a range of key issues in the field, many of which are unresolved Received in revised form and some of which are controversial. (1) El Nino-Southern~ Oscillation variability, while the most 8 June 2015 important global-scale mode of interannual climate variation, is insufficient to explain most of the Accepted 10 June 2015 variation of tropical South American climate observed in both the instrumental and the paleoclimate Available online xxx records. (2) Significant climate variation in tropical South America occurs on seasonal to orbital (i.e. multi-millennial) time scales as a result of sea-surface temperature (SST) variation and ocean Keywords: e Paleoclimate atmosphere interactions of the tropical Atlantic. (3) Decadal-scale climate variability, linked with this Monsoon tropical Atlantic variability, has been a persistent characteristic of climate in tropical South America for at Amazon least the past half millennium, and likely, far beyond. -
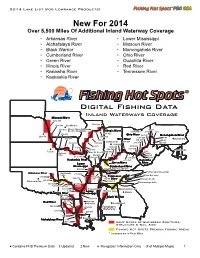
2014 Lake List (For Lowrance Products)
2014 Lake List (for Lowrance Products) New For 2014 Over 5,500 Miles Of Additional Inland Waterway Coverage • Arkansas River • Lower Mississippi • Atchafalaya River • Missouri River • Black Warrior • Monongahela River • Cumberland River • Ohio River • Green River • Ouachita River • Illinois River • Red River • Kanawha River • Tennessee River • Kaskaskia River Digital Fishing Data Inland Waterways Coverage Missouri River 600-750 Chicago 500-600 Des Moines 400-500 Illinois River (Starved 11-31 Rock to Hennepin) Illinois River Cleveland 31-86 1-11 300-400 301-322 Ohio River 86-127 Monongahela River Lincoln 256-301 1-41 200-300 127-165 Harrisburg 231-256 Ohio River 165-203 41-86 Indianapolis 438-534 100-200 137-206 203-274 86-128 534-582 82-137 274-311 1-100 582-644 391-438 21-82 644-735 Annapolis Topeka Miss. R. Pool 26 735-852 1-24 Kansas 24-49 City 852-886 St. Louis 49-65 886-981 311-331 65-91 Kaskaskia River 331-391 Kanawha Lower Green River River Mississippi Cumberland River 828-950 93-148 270-381 148-220 Fort Loudon Reservoir Arkansas River Dardanelle 716-828 Lake Watts Bar Lake 371-444 Kentucky Tennessee 600-716 Lake 133-68 River Nickajack Lake 330-371 133-214 Oklahoma City Chickamauga Lake 262-330 Little 197-262 Rock 126-197 61-127 Pickwick Lake Guntersville Lake 1-65 Lower Wheeler Lake Ouachita River Mississippi Atlanta 255-337 Black Warrior 177-255 520-600 Red River 89-177 425-520 Jackson Montgomery 5-89 166-237 325-425 Tombigbee Waterway 73-166 236-325 73-6 Baton Rouge Atchafalaya River 1-46 46-118 Army Corps of Engineers Contours, Structure & Nav. -

Download The
SYSTEMATICS OF SJ^IFRJGA BOFIDULJ AND BELATED SPECIES FBCM THE COLOMBIA BIVEB GOBGE TO SOUTHSESTEBN BBITISH COIUUBIA By HALTER ETHEN PERKINS B.Sc. , University of Oklahoma, 1S73 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOB THE DEGREE OF DOCTOB OF PHILOSOPHY in THE FACULTY OF GRADUATE STUDIES {Department of Botany) Se accept this thesis as conforming to the required standard THE UNIVERSITY OF BBITISH COLUMBIA October, 1978 (8) Walter Ethen Perkins, 1978 In presenting this thesis in partial fulfilment of the requirements for an advanced degree at the University of British Columbia, I agree that the Library shall make it freely available for reference and study. I further agree that permission for extensive copying of this thesis for scholarly purposes may be granted by the Head of my Department or by his representatives. It is understood that copying or publication of this thesis for financial gain shall not be allowed without my written permission. W. Ethen Perkins Department of Botany The University of British Columbia 2075 Wesbrook Place Vancouver, Canada V6T 1W5 5 October, 1978 Research Supervisor: Dr. Ratherine I. Beamish. ABSTRACT In the Pacific Northwest, hybridization and polypoid variation have produced confusion in the relationships among Saxifraga rufidula (Small)James Macoun and its relatives. The entities from the Columbia Biver Gorge are particularly difficult to separate taxoncmically., Some authors have recognized several species from that area while others recognize one highly variable, widely distributed species with varietal components. The present study approached the systematic treatment of S. rufidula and related subspecif ic taxa of S. occidentalis Wats. -

The Ice Age Revealed
Warwickshire Geological The Big Lake? Our Work Conservation Group It is thought that during the glaciation, Today the sands and gravels are extracted as a source of the ice sheets blocked the Bytham River aggregate for the production of concrete and mortar used in the to form a large glacial lake or lakes that construction industry. With backing from the Aggregates Levy some geologists have referred to as Sustainability Fund through English Nature and support from the ‘Lake Harrison’. Finely banded clays University of Warwick, Warwickshire Museum and Smith’s seen in local quarries might represent Concrete, WGCG are running a project to interpret and conserve The Ice Age ancient lake-bottom mud. the sand, gravel and clay deposits found in eastern Warwickshire. An interpretation board has been installed at Ryton Pools Country Park, Bubbenhall and exposures of ancient Revealed river and glacial deposits have been protected at the country Coventry park and at Wood Farm Quarry. Look out for more information Bubbenhall around local villages and at the Wildlife Trust’s Brandon Marsh Ryton Pools Ryton Wood Country Park Nature Centre, Brandon. A445 Warwickshire Geological Conservation Group is a non-profit Leamington Spa Wood Farm Quarry making organisation interested in promoting and protecting local Bubbenhall Wood earth heritage within local communities. If you would like to Wappenbury obtain more information or become a member and get involved, Wood please contact us: Warwickshire Geological Conservation Group c/o Centre for Lifelong Learning University of Warwick Coventry CV4 7AL Telephone 02476 52 35 33 Baggington sand www.wgcg.co.uk www.woodfarmproject.org.uk Baggington gravel The Ice Age Revealed Parts of eastern Warwickshire are Warwickshire’s First People The Big Chill underlain by deposits of sand, gravel The first people to colonise Britain Around 450,000 years ago Britain and clay. -

History of Bass Stocking in Ontario
Bass Stocking and Transfers: An Annotated Bibliography and Literature Review T. A. Lasenby and S. J. Kerr Fisheries Section Fish and Wildlife Branch Ontario Ministry of Natural Resources December 2000 This publication should be cited as follows: Lasenby, T. A. and S. J. Kerr. 2000. Bass transfers and stocking: An annotated bibliography and literature review. Fish and Wildlife Branch, Ontario Ministry of Natural Resources. Peterborough, Ontario. 207 p. + appendices. Printed in Ontario, Canada (0.3 k P. R. 00 15 12) MNR 51461 ISBN 0-7794-0488-2 Copies of this publication are available from: Fish and Wildlife Branch Ontario Ministry of Natural Resources P. O. Box 7000 300 Water Street Peterborough, Ontario K9J 8M5 CetteCette publication publication spécialisée spécialisée n’est n’ estdisponible disponible qu’en qu’en anglais angl Cover drawing by Ruth E. Grant, Brockville, Ontario. Preface This bibliography and literature review is the fourth in a series of reference documents developed in conjunction with a review of fish stocking policies and guidelines for the Province of Ontario. It has been prepared to summarize information on the current state of knowledge regarding the stocking and transfer of bass (smallmouth and largemouth) in a form which can be readily utilized by field staff and stocking proponents. Material cited in this bibliography includes papers published in scientific journals, magazines and periodicals as well as “gray” literature such as file reports from the Ministry of Natural Resources (MNR) field offices. Unpublished literature was obtained by soliciting information (i.e., unpublished data and file reports) from field biologists from across Ontario. -

IA Alaskaexp13harruoft
iumi !!!!!iO,'!t':;:!i;'^i-'^!!'!|! \iM.m'' m- mMmmm:' i =m iiiSlii!-!'' mm}^\ mm': !|lilltinilililiHtnt)Uili)Uillti)iiiiti>lliiiillllillltltitll Digitized by the Internet Archive in 2007 with funding from IVIicrosoft Corporation http://www.archive.org/details/alaskaexp13harruoft ALASKA VOLUME xm I Ife^^^ SMITHSONIAN INSTITUTION HARRIMAN ALASKA SERIES VOLUME XIII LAND AND FRESH WATER MOLLUSKS BY WILLIAM H. DALL HYDROIDS BY C. C. NUTTING (Publication 2000) CITY OF WASHINGTON PUBLISHED BY THE SMITHSONIAN INSTITUTION 1910 Q 115 V. lb ADVERTISEMENT. The publication of the series of volumes on the Harriman Alaska Expedition of 1899, heretofore pri- vately printed, has been transferred to the Smithsonian Institution by Mrs. Edward H. Harriman, and the work will hereafter be known as the Harriman Alaska Series of the Smithsonian Institution. The remainder of the edition of Volumes I to V, and VIII to XIII, as also Volumes VI and VII in preparation, together with any additional volumes that may hereafter appear, will bear special Smithsonian title pages. Smithsonian Institution, Washington, D. C, July, 1910 HARRIMAN ALASKA EXPEDITION WITH COOPERATION OF WASHINGTON ACADEMY OF SCIENCES ALASKA VOLUME XIII LAND AND FRESH WATER MOLLUSKS BY WILLIAM H. DALL HYDROIDS BY C. C. NUTTING NEW YORK DOUBLEDAY. PAGE & COMPANY 1905 Copyright, 1905 BY Edward H. Harriman ; PREFACE The present volume comprises two papers : one on the Land and Fresh Water Mollusks of Alaska, by Dr. Wm. H. Dall the other on the Hydroids of the Expedition, by Prof. C. C. Nutting. Dr. DalFs paper has not been previously published Professor Nutting's appeared in the Proceedings of the Wash- ington Academy of Sciences in May, 1901.