Flavor Chemistry of Butter Culture
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Flavour Management by Citric Acid Negative MLF Starter Cultures Authors: Carsten Heinemeyer, 2B Fermcontrol; Ulrich Hamm DLR-RNH Bad Kreuznach, Germany; Dr
Flavour management by citric acid negative MLF starter cultures Authors: Carsten Heinemeyer, 2B FermControl; Ulrich Hamm DLR-RNH Bad Kreuznach, Germany; Dr. Jürgen Fröhlich, University of Mainz General will be completed in all fermentations. Additionally, acetic acid and a certain residual profile of the metabolic intermediates The malo-lactic fermentation (MLF) is a commonly used remain in the wine. method to convert the aggressive malic acid to lactic acid. This conversion results in a reduction of the titratable acidity, which New MLF Starter Cultures is desired mainly in red wine but also in numerous white wines. The development of a citric acid negative MLF starter culture This process will be done either by the indigenous flora of LAB gave a new opportunity to avoid diacetyl and acetic acid or by selected strains of LAB e.g. Oenococcus oeni. During the production from the citric acid degradation. The fermentation MLF Oenococcus oeni does not convert only malic acid into lactic with a citric acid negative MLF starter culture will best preserve acid, numerous amounts of aroma active by-products will also be the varietal character of the wine. produced. The best known is diacetyl, which gives buttery notes to the wine. Diacetyl will be produced during the MLF by the In a comprehensive four year study at the wine research institute conversion of the natural citric acid in the wine by Oenococcus DLR-RNH Bad Kreuznach in Rhineland-Palatinate-Germany, oeni (Jan Clair Nielsen 1999). Apart from the diacetyl formation different MLF starter strains were tested under practical by Oenococcus oeni, numerous intermediate by-products are winemaking conditions. -

Acetoin Acetate Natural
aurochemicals.com Safety Data Sheet HEALTH 0 FLAMMABILITY 2 REACTIVITY 0 Section 1: PRODUCT AND COMPANY IDENTIFICATION 1.1 Product identifiers Product Name Acetoin Acetate, Natural Product Number 0352600 CAS-No. 4906-24-5 1.2 Product Recommended Use Flavorings 1.3 Preparation Information Company Aurochemicals 7 Nicoll Street Washingtonville, NY 10992- USA Telephone 845-496-6065 Fax 845-496-6248 1.4 Emergency Telephone Number 1-800-535-5053 International - 1-352-323-3500 collect Section 2: HAZARD(s) IDENTIFICATION 2.1 Classification of substance or mixture GHS Classification in accordance with 29 CFR 1910 (OSHA HCS) Combustible Liquid (Category 4) H227 Skin irritation (Category 2) H315 Eye Irritation (Category 2A) H319 Specific Target Organ Toxicity -Single Exposure -Respiratory irritation H335 2.2 GHS Label Elements, Including precautionary statements Pictogram Signal Statement Warning Hazard Statement(s) H227 Combustible liquid H315 Causes skin irritation H319 Causes serious eye irritation H335 May cause respiratory irritation Precautionary Statement(s) PREVENTION P210 Keep away from flames and hot surfaces - No smoking P216 Avoid breathing dust/fumes/gas/mist/vapors/spray P280 Wear protective gloves/protective clothing/face protection 3526 ACETOIN ACETATE Nat sds.doc Printed: April 17, 2019 Page 1 of 7 aurochemicals.com Safety Data Sheet RESPONSE P305+351+338 IF IN EYES: Rinse cautiously with water for several minutes. Remove contact lenses if present and easy to do. Continue rinsing. STORAGE P405 Store locked up DISPOSAL P501 Dispose of contents/container to an approved waste disposal plant 2.3 HNOC (Hazards not otherwise None classified or not covered by GHS Section 3: COMPOSITION / INFORMATION ON INGREDIENTS 3.1 Substances Synonym 2-Acetoxy-3-Butanone Formula C6H10O3 Molecular Weight 130.14 g/mol CAS-No 4906-24-5 EC-No 200-580-7 Hazardous Components Does not contain any hazardous substances Section 4: FIRST AID MEASURES 4.1 Description of first aid measures General Advice Consult a physician. -

Retention Indices for Frequently Reported Compounds of Plant Essential Oils
Retention Indices for Frequently Reported Compounds of Plant Essential Oils V. I. Babushok,a) P. J. Linstrom, and I. G. Zenkevichb) National Institute of Standards and Technology, Gaithersburg, Maryland 20899, USA (Received 1 August 2011; accepted 27 September 2011; published online 29 November 2011) Gas chromatographic retention indices were evaluated for 505 frequently reported plant essential oil components using a large retention index database. Retention data are presented for three types of commonly used stationary phases: dimethyl silicone (nonpolar), dimethyl sili- cone with 5% phenyl groups (slightly polar), and polyethylene glycol (polar) stationary phases. The evaluations are based on the treatment of multiple measurements with the number of data records ranging from about 5 to 800 per compound. Data analysis was limited to temperature programmed conditions. The data reported include the average and median values of retention index with standard deviations and confidence intervals. VC 2011 by the U.S. Secretary of Commerce on behalf of the United States. All rights reserved. [doi:10.1063/1.3653552] Key words: essential oils; gas chromatography; Kova´ts indices; linear indices; retention indices; identification; flavor; olfaction. CONTENTS 1. Introduction The practical applications of plant essential oils are very 1. Introduction................................ 1 diverse. They are used for the production of food, drugs, per- fumes, aromatherapy, and many other applications.1–4 The 2. Retention Indices ........................... 2 need for identification of essential oil components ranges 3. Retention Data Presentation and Discussion . 2 from product quality control to basic research. The identifi- 4. Summary.................................. 45 cation of unknown compounds remains a complex problem, in spite of great progress made in analytical techniques over 5. -

Sauces Reconsidered
SAUCES RECONSIDERED Rowman & Littlefield Studies in Food and Gastronomy General Editor: Ken Albala, Professor of History, University of the Pacific ([email protected]) Rowman & Littlefield Executive Editor: Suzanne Staszak-Silva ([email protected]) Food studies is a vibrant and thriving field encompassing not only cooking and eating habits but also issues such as health, sustainability, food safety, and animal rights. Scholars in disciplines as diverse as history, anthropol- ogy, sociology, literature, and the arts focus on food. The mission of Row- man & Littlefield Studies in Food and Gastronomy is to publish the best in food scholarship, harnessing the energy, ideas, and creativity of a wide array of food writers today. This broad line of food-related titles will range from food history, interdisciplinary food studies monographs, general inter- est series, and popular trade titles to textbooks for students and budding chefs, scholarly cookbooks, and reference works. Appetites and Aspirations in Vietnam: Food and Drink in the Long Nine- teenth Century, by Erica J. Peters Three World Cuisines: Italian, Mexican, Chinese, by Ken Albala Food and Social Media: You Are What You Tweet, by Signe Rousseau Food and the Novel in Nineteenth-Century America, by Mark McWilliams Man Bites Dog: Hot Dog Culture in America, by Bruce Kraig and Patty Carroll A Year in Food and Beer: Recipes and Beer Pairings for Every Season, by Emily Baime and Darin Michaels Celebraciones Mexicanas: History, Traditions, and Recipes, by Andrea Law- son Gray and Adriana Almazán Lahl The Food Section: Newspaper Women and the Culinary Community, by Kimberly Wilmot Voss Small Batch: Pickles, Cheese, Chocolate, Spirits, and the Return of Artisanal Foods, by Suzanne Cope Food History Almanac: Over 1,300 Years of World Culinary History, Cul- ture, and Social Influence, by Janet Clarkson Cooking and Eating in Renaissance Italy: From Kitchen to Table, by Kath- erine A. -

Some of the Factors Influencing the Growth of Molds in Butter Harold Macy Iowa State College
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1929 Some of the factors influencing the growth of molds in butter Harold Macy Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Agriculture Commons, and the Food Science Commons Recommended Citation Macy, Harold, "Some of the factors influencing the growth of molds in butter " (1929). Retrospective Theses and Dissertations. 14244. https://lib.dr.iastate.edu/rtd/14244 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the miaofilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. in the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Al.so, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections vwth small overiaps. -

Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review
fermentation Review Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review Valentina Martin, Maria Jose Valera , Karina Medina, Eduardo Boido and Francisco Carrau * Enology and Fermentation Biotechnology Area, Food Science and Technology Department, Facultad de Quimica, Universidad de la Republica, Montevideo 11800, Uruguay; [email protected] (V.M.); [email protected] (M.J.V.); [email protected] (K.M.); [email protected] (E.B.) * Correspondence: [email protected]; Tel.: +598-292-481-94 Received: 7 August 2018; Accepted: 5 September 2018; Published: 10 September 2018 Abstract: Apiculate yeasts of the genus Hanseniaspora/Kloeckera are the main species present on mature grapes and play a significant role at the beginning of fermentation, producing enzymes and aroma compounds that expand the diversity of wine color and flavor. Ten species of the genus Hanseniaspora have been recovered from grapes and are associated in two groups: H. valbyensis, H. guilliermondii, H. uvarum, H. opuntiae, H. thailandica, H. meyeri, and H. clermontiae; and H. vineae, H. osmophila, and H. occidentalis. This review focuses on the application of some strains belonging to this genus in co-fermentation with Saccharomyces cerevisiae that demonstrates their positive contribution to winemaking. Some consistent results have shown more intense flavors and complex, full-bodied wines, compared with wines produced by the use of S. cerevisiae alone. Recent genetic and physiologic studies have improved the knowledge of the Hanseniaspora/Kloeckera species. Significant increases in acetyl esters, benzenoids, and sesquiterpene flavor compounds, and relative decreases in alcohols and acids have been reported, due to different fermentation pathways compared to conventional wine yeasts. -

Breakfast Glycaemic Response in Patients with Type 2 Diabetes: Effects of Bedtime Dietary Carbohydrates
European Journal of Clinical Nutrition (1999) 53, 706±710 ß 1999 Stockton Press. All rights reserved 0954±3007/99 $15.00 http://www.stockton-press.co.uk/ejcn Breakfast glycaemic response in patients with type 2 diabetes: Effects of bedtime dietary carbohydrates M Axelsen1*, R Arvidsson Lenner 2,PLoÈnnroth1 and U Smith 1 1The Lundberg Laboratory for Diabetes Research, Department of Internal Medicine, Sahlgrenska University Hospital, S-413 45 GoÈteborg, Sweden; and 2Department of Clinical Nutrition, Department of Internal Medicine, Sahlgrenska University Hospital, GoÈteborg University, Sweden Objectives: Bedtime carbohydrate (CHO) intake in patients with type-2 diabetes may improve glucose tolerance at breakfast the next morning. We examined the `overnight second-meal effect' of bedtime supplements containing `rapid' or `slow' CHOs. Design: Randomized cross-over study with three test-periods, each consisting of two days on a standardized diet, followed by a breakfast tolerance test on the third morning. Setting: The Lundberg Laboratory for Diabetes Research, Sahlgrenska University Hospital, GoÈteborg, Sweden. Subjects: Sixteen patients with type 2 diabetes on oral agents and=or diet. Interventions: Two different bedtime (22.00 h) CHO supplements (0.46 g available CHO=kg body weight) were compared to a starch-free placebo (`normal' food regimen). The CHOs were provided as uncooked cornstarch (slow-release CHOs) or white bread (rapid CHOs). Results: On the mornings after different bedtime meals we found similar fasting glucose, insulin, free fatty acid and lactate levels. However, the glycaemic response after breakfast was 21% less after uncooked cornstarch compared to placebo ingestion at bedtime (406Æ 46 vs 511Æ 61 mmol min 171, P < 0.01). -

An Original Method for Producing Acetaldehyde and Diacetyl by Yeast
b r a z i l i a n j o u r n a l o f m i c r o b i o l o g y 4 7 (2 0 1 6) 949–954 ht tp://www.bjmicrobiol.com.br/ Industrial Microbiology An original method for producing acetaldehyde and diacetyl by yeast fermentation a a,∗ a b a,c Irina Rosca , Anca Roxana Petrovici , Mihai Brebu , Irina Stoica , Bogdan Minea , a Narcisa Marangoci a “Petru Poni” Institute of Macromolecular Chemistry, Advanced Research Center for Bionanoconjugates and Biopolymers, Aleea GrigoreGhica Voda, Iasi, Romania b SC Zeelandia SRL, R&D Department, Valea Lupului, Iasi, Romania c “Grigore T. Popa” University of Medicine and Pharmacy, Iasi, Romania a r a t i c l e i n f o b s t r a c t Article history: In this study a natural culture medium that mimics the synthetic yeast peptone glucose Received 2 June 2015 medium used for yeast fermentations was designed to screen and select yeasts capable Accepted 29 February 2016 of producing high levels of diacetyl and acetaldehyde. The presence of whey powder and Available online 25 July 2016 sodium citrate in the medium along with manganese and magnesium sulfate enhanced Associate Editor: Jorge Gonzalo both biomass and aroma development. A total of 52 yeasts strains were cultivated in two Farias Avendano different culture media, namely, yeast peptone glucose medium and yeast acetaldehyde- diacetyl medium. The initial screening of the strains was based on the qualitative reaction Keywords: of the acetaldehyde with Schiff’s reagent (violet color) and diacetyl with Brady’s reagent Acetaldehyde (yellow precipitate). -

An Unrecognized Hazard in E-Cigarette Vapor: Preliminary Quantification of Methylglyoxal Formation from Propylene Glycol in E-Cigarettes
International Journal of Environmental Research and Public Health Article An Unrecognized Hazard in E-Cigarette Vapor: Preliminary Quantification of Methylglyoxal Formation from Propylene Glycol in E-Cigarettes Parham Azimi 1, Zahra Keshavarz 1, Marianne Lahaie Luna 1,2, Jose Guillermo Cedeno Laurent 1 , Jose Vallarino 1, David C. Christiani 1 and Joseph G. Allen 1,* 1 Department of Environmental Health, Harvard T. H. Chan School of Public Health, Boston, MA 02115, USA; [email protected] (P.A.); [email protected] (Z.K.); [email protected] (M.L.L.); [email protected] (J.G.C.L.); [email protected] (J.V.); [email protected] (D.C.C.) 2 Occupational & Environmental Health Division, Dalla Lana School of Public Health, University of Toronto, Toronto, ON M5T 3M7, Canada * Correspondence: [email protected] Abstract: Up to 95% of the liquid volume in an e-cigarette consists of propylene glycol. Previous research has shown that propylene glycol can generate diacetyl and formaldehyde when heated. New research shows that propylene glycol can also generate methylglyoxal, an alpha di-carbonyl compound recently shown to cause epithelial necrosis at even lower concentrations than diacetyl, the flavoring chemical associated with bronchiolitis obliterans (“Popcorn Lung”). We analyzed chemical emissions from 13 JUUL pod flavors. Diacetyl and methylglyoxal was detected in 100% of samples 3 3 with median concentration (range) of 20 µg/m (less than limit of quantification: 54 µg/m ) and 4219 µg/m3 (677–15,342 µg/m3), respectively. We also detected acetaldehyde (median concentration: Citation: Azimi, P.; Keshavarz, Z.; 341 µg/m3) and propionaldehyde (median concentration: 87 µg/m3) in all samples. -

Developing the Butter Value Chain in Ethiopia
LIVES WORKING PAPER 1 PAPER WORKING LIVES Developing the butter value chain in Ethiopia ISBN 92–9146–366–3 Livestock and irrigation value chains for Ethiopian smallholders project aims to improve the competitiveness, sustainability and equity of value chains for selected high‐value livestock and irrigated crop commodities in target areas of four regions of Ethiopia. It identifies, targets and promotes improved technologies and innovations to develop high value livestock and irrigated crop value chains; it improves the capacities of Livestock and Irrigation Value chains for Ethiopian Smallholders value chain actors; it improves the use of knowledge at different levels; it generates knowledge through action‐oriented research; and it promotes and disseminates good practices. Project carried out with the financial support of the Government of Canada provided through Foreign Affairs, Trade and Development Canada (DFATD). lives-ethiopia.org The International Livestock Research Institute (ILRI) works to improve food security and reduce poverty in developing countries through research for better and more sustainable use of livestock. ILRI is a member of the CGIAR Consortium, a global research partnership of 15 centres working with many partners for a food-secure future. ILRI has two main campuses in East Africa and other hubs in East, West and southern Africa and South, Southeast and East Asia. ilri.org The International Water Management Institute (IWMI) is a non-profit, scientific research organization focusing on the sustainable use of water and land resources in developing countries. It is headquartered in Colombo, Sri Lanka, with regional offices across Asia and Africa. IWMI works in partnership with governments, civil society and the private sector to develop scalable agricultural water management solutions that have a real impact on poverty reduction, food security and ecosystem health. -

Diabetes Exchange List
THE DIABETIC EXCHANGE LIST (EXCHANGE DIET) The Exchange Lists are the basis of a meal planning system designed by a committee of the American Diabetes Association and the American Dietetic Association. The Exchange Lists The reason for dividing food into six different groups is that foods vary in their carbohydrate, protein, fat, and calorie content. Each exchange list contains foods that are alike; each food choice on a list contains about the same amount of carbohydrate, protein, fat, and calories as the other choices on that list. The following chart shows the amounts of nutrients in one serving from each exchange list. As you read the exchange lists, you will notice that one choice is often a larger amount of food than another choice from the same list. Because foods are so different, each food is measured or weighed so that the amounts of carbohydrate, protein, fat, and calories are the same in each choice. The Diabetic Exchange List Carbohydrate (grams) Protein (grams) Fat (grams) Calories I. Starch/Bread 15 3 trace 80 II. Meat Very Lean - 7 0-1 35 Lean - 7 3 55 Medium-Fat - 7 5 75 High-Fat - 7 8 100 III. Vegetable 5 2 - 25 IV. Fruit 15 - - 60 V. Milk Skim 12 8 0-3 90 Low-fat 12 8 5 120 Whole 12 8 8 150 VI. Fat - - 5 45 You will notice symbols on some foods in the exchange groups. 1. Foods that are high in fiber (three grams or more per normal serving) have the symbol *. 2. Foods that are high in sodium (400 milligrams or more of sodium per normal serving) have the symbol #. -
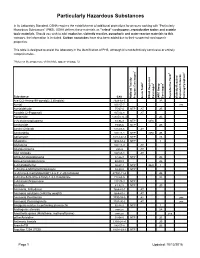
Particularly Hazardous Substances
Particularly Hazardous Substances In its Laboratory Standard, OSHA requires the establishment of additional protections for persons working with "Particularly Hazardous Substances" (PHS). OSHA defines these materials as "select" carcinogens, reproductive toxins and acutely toxic materials. Should you wish to add: explosive, violently reactive, pyrophoric and water-reactve materials to this category, the information is included. Carbon nanotubes have also been added due to their suspected carcinogenic properties. This table is designed to assist the laboratory in the identification of PHS, although it is not definitively conclusive or entirely comprehensive. *Notes on the proper use of this table appear on page 12. 1 6 5 2 3 4 Substance CAS National Toxicity National Program Carcinogen Toxin Acute Regulated OSHA Carcinogen Group IARC Carcinogen Toxin Reproductive Violently Reactive/ Explosive/Peroxide Forming/Pyrophoric A-a-C(2-Amino-9H-pyrido[2,3,b]indole) 2648-68-5 2B Acetal 105-57-7 yes Acetaldehyde 75-07-0 NTP AT 2B Acrolein (2-Propenal) 107-02-8 AT Acetamide 126850-14-4 2B 2-Acetylaminofluorene 53-96-3 NTP ORC Acrylamide 79-06-6 NTP 2B Acrylyl Chloride 814-68-6 AT Acrylonitrile 107-13-1 NTP ORC 2B Adriamycin 23214-92-8 NTP 2A Aflatoxins 1402-68-2 NTP 1 Allylamine 107-11-9 AT Alkylaluminums varies AT Allyl Chloride 107-05-1 AT ortho-Aminoazotoluene 97-56-3 NTP 2B para-aminoazobenzene 60-09-3 2B 4-Aminobiphenyl 92-67-1 NTP ORC 1 1-Amino-2-Methylanthraquinone 82-28-0 NTP (2-Amino-6-methyldipyrido[1,2-a:3’,2’-d]imidazole) 67730-11-4 2B