State of the Physical, Biological and Selected Fishery Resources of Pacific Canadian Marine Ecosystems in 2017
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
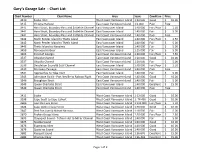
Gary's Charts
Gary’s Garage Sale - Chart List Chart Number Chart Name Area Scale Condition Price 3410 Sooke Inlet West Coast Vancouver Island 1:20 000 Good $ 10.00 3415 Victoria Harbour East Coast Vancouver Island 1:6 000 Poor Free 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair $ 5.00 3441 Haro Strait, Boundary Pass and Sattelite Channel East Coast Vancouver Island 1:40 000 Poor Free 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair $ 5.00 3443 Thetis Island to Nanaimo East Vancouver Island 1:40 000 Fair $ 5.00 3459 Nanoose Harbour East Vancouver Island 1:15 000 Fair $ 5.00 3463 Strait of Georgia East Coast Vancouver Island 1:40 000 Fair/Poor $ 7.50 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Good $ 10.00 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Fair $ 5.00 3538 Desolation Sound & Sutil Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3539 Discovery Passage East Coast Vancouver Island 1:40 000 Poor Free 3541 Approaches to Toba Inlet East Vancouver Island 1:40 000 Fair $ 5.00 3545 Johnstone Strait - Port Neville to Robson Bight East Coast Vancouver Island 1:40 000 Good $ 10.00 3546 Broughton Strait East Coast Vancouver Island 1:40 000 Fair $ 5.00 3549 Queen Charlotte Strait East Vancouver Island 1:40 000 Excellent $ 15.00 3549 Queen Charlotte Strait East -

Color Patterns of the Shrimps Heptacarpus Pictus and H. Paludicola (Caridea: Hippolytidae)
MARINE BIOLOGY Marine Biology 64, 141-152 (1981) © Springer-Verlag 1981 Color Patterns of the Shrimps Heptacarpus pictus and H. paludicola (Caridea: Hippolytidae) R. T. Bauer Department of Invertebrate Zoology, National Museum of Natural History, Smithsonian Institution; Washington, D.C. 20560, USA Abstract coloration has been on the isopod Idothea montereyensis Maloney; Lee (1966 a,b, 1972) studied the biochemical Color patterns of the shallow-water shrimps Heptacarpus and cellular bases of color patterns and the ecology of pictus and H. paludicola are formed by chromatosomes color change. However, no caridean shrimp has been (usually termed chromatophores) located beneath the studied so thoroughly. Gamble and Keeble (1900) and translucent exoskeleton. Development of color patterns Keeble and Gamble (1900, 1904) described in detail the is related to size (age) and sex. The color expressed is chromatophores (chromatosomes in the modern usage of determined by the chromatosome pigment dispersion, Elofsson and Kauri, 1971) and coloration of the shrimp arrangement, and density. In populations with well- Hippolyte varians Leach. Chassard-Bouchaud (1965), in developed coloration (//. pictus from Cayucos, California, her extensive work on physiological and morphological 1976-1978,//. paludicola from ArgyleChannel, San Juan color change in caridean shrimp, described the chromato Island, Washington, June-July, 1978), prominent somes and illustrated the color morphs of H. varians, coloration was a characteristic of maturing females, Palaemoii squilla (L.), P. serratus (Pennant), Athanas breeding females, and some of the larger males. In the nitescens Leach, and Crangon crangon (L.). Carlisle and Morro Bay, California, population of H. paludicola Knowles (1959) give a brief review of color patterns in (sampled 1976-1978), color patterns were poorly decapod species. -

Sagitta Siamensis, a New Benthoplanktonic Chaetognatha Living in Marine Meadows of the Andaman Sea, Thailand
Cah. Biol. Mar. (1997) 38 : 51-58 Sagitta siamensis, a new benthoplanktonic Chaetognatha living in marine meadows of the Andaman Sea, Thailand Jean-Paul Casanova (1) and Taichiro Goto(2) (1) Laboratoire de Biologie animale (Plancton), Université de Provence, Place Victor Hugo, 13331 Marseille Cedex 3, France Fax : (33) 4 91 10 60 06; E-mail: [email protected] (2) Department of Biology, Faculty of Education, Mie University, Tsu, Mie 514, Japan Abstract: A new benthoplanktonic chaetognath, Sagitta siamensis, is described from near-shore waters of Phuket Island (Thailand), in the Andaman Sea, where it lives among submerged vegetation. It is related to the species of the "hispida" group. In the laboratory, specimens have been observed swimming in the sea water but also sometimes adhering to the wall of the jars, and the eggs are benthic and attached on the substratum. Their fins are particularly thick and provided with clusters of probably adhesive cells on their ventral side and edges. This is the first mention of such fins in the genus Sagitta but the adhesive apparatus do not resemble that found in the benthic family Spadellidae and is less evolved. A review of the morphological characteristics of the species of the "hispida" group is done as well as their biogeography. Résumé : Un nouveau chaetognathe benthoplanctonique, Sagitta siamensis, est décrit des eaux néritiques de l'île Phuket (Thaïlande) en mer des Andaman, où il vit dans la végétation immergée. Il est proche des espèces du groupe "hispida". Au Laboratoire, les spécimens ont été observés nageant dans l'eau mais parfois aussi adhérant aux parois des récipients et les œufs sont benthiques, fixés sur le substratum. -

Gwaii Haanas National Park Reserve and Haida Heritage Site
GWAII HAANAS NATIONAL PARK RESERVE AND HAIDA HERITAGE SITE Management Plan for the Terrestrial Area A Pacific coast wilderness in Haida Gwaii — the Queen Charlotte Islands. Protected through the cooperation of the Government of Canada and the Council of the Haida Nation Produced by the ARCHIPELAGO MANAGEMENT BOARD GWAII HAANAS STRATEGIC MANAGEMENT PLAN FOR THE TERRESTRIAL AREA FOREWORD n January of 1993, the Government of The Council of the Haida Nation and the I Canada and the Council of the Haida Nation Government of Canada agree and support the signed the Gwaii Haanas Agreement. In this contents of this plan and will work together document, both parties stated their commitment through the Archipelago Management Board to the protection of Gwaii Haanas, one of the to implement the plan’s recommendations. world’s great natural and cultural treasures. A By supporting this plan, the two parties part of this agreement describes the cooperative assert their belief in the value and benefit of management procedures, including establish- cooperative management and preservation of ment of the Archipelago Management Board. Gwaii Haanas. This management plan, produced by the Archipelago Management Board in consultation with the public, sets out strategic objectives Approved by: for appropriate use and protection of Gwaii Haanas. The plan not only provides comprehensive strategic direction for managing ○○○○○○○○○○○○○ Gwaii Haanas, but it also serves as an example ○○○○○○○○○○○○○○○○ for the Government of Canada of cooperative effort and marks an important milestone in the relationships of Canada and the Haida Nation. ○○○○○○○○○○○○○○○○○○○○○○○○○○○○○ for the Council of the Haida Nation 3i CONTENTS 1 INTRODUCTION ....................................................................................... 2 1.1 Description of Gwaii Haanas......................................................... -

The Histology of Nanomia Bijuga (Hydrozoa: Siphonophora) SAMUEL H
RESEARCH ARTICLE The Histology of Nanomia bijuga (Hydrozoa: Siphonophora) SAMUEL H. CHURCH*, STEFAN SIEBERT, PATHIKRIT BHATTACHARYYA, AND CASEY W. DUNN Department of Ecology and Evolutionary Biology, Brown University, Providence, Rhode Island ABSTRACT The siphonophore Nanomia bijuga is a pelagic hydrozoan (Cnidaria) with complex morphological organization. Each siphonophore is made up of many asexually produced, genetically identical zooids that are functionally specialized and morphologically distinct. These zooids predominantly arise by budding in two growth zones, and are arranged in precise patterns. This study describes the cellular anatomy of several zooid types, the stem, and the gas-filled float, called the pneumatophore. The distribution of cellular morphologies across zooid types enhances our understanding of zooid function. The unique absorptive cells in the palpon, for example, indicate specialized intracellular digestive processing in this zooid type. Though cnidarians are usually thought of as mono-epithelial, we characterize at least two cellular populations in this species which are not connected to a basement membrane. This work provides a greater understanding of epithelial diversity within the cnidarians, and will be a foundation for future studies on N. bijuga, including functional assays and gene expression analyses. J. Exp. Zool. (Mol. Dev. Evol.) 324B:435– 449, 2015. © 2015 The Authors. Journal of Experimental Zoology Part B: Molecular and J. Exp. Zool. Developmental Evolution Published by Wiley Periodicals, Inc. (Mol. Dev. Evol.) 324B:435–449, How to cite this article: Church SH, Siebert S, Bhattacharyya P, Dunn CW. 2015. The histology of 2015 Nanomia bijuga (Hydrozoa: Siphonophora). J. Exp. Zool. (Mol. Dev. Evol.) 324B:435–449. Siphonophores are pelagic hydrozoans (Cnidaria) with a highly and defense (Dunn and Wagner, 2006; Totton, '65). -

Demography and the Evolution of Logistic Organization on the Northern Northwest Coast Between 11,000 and 5,000 Cal BP
Portland State University PDXScholar Dissertations and Theses Dissertations and Theses Winter 7-20-2016 Demography and the Evolution of Logistic Organization on the Northern Northwest Coast Between 11,000 and 5,000 cal BP Thomas Jay Brown Portland State University Follow this and additional works at: https://pdxscholar.library.pdx.edu/open_access_etds Part of the Anthropology Commons Let us know how access to this document benefits ou.y Recommended Citation Brown, Thomas Jay, "Demography and the Evolution of Logistic Organization on the Northern Northwest Coast Between 11,000 and 5,000 cal BP" (2016). Dissertations and Theses. Paper 3223. https://doi.org/10.15760/etd.3218 This Thesis is brought to you for free and open access. It has been accepted for inclusion in Dissertations and Theses by an authorized administrator of PDXScholar. Please contact us if we can make this document more accessible: [email protected]. Demography and the Evolution of Logistic Organization on the Northern Northwest Coast Between 11,000 and 5,000 cal BP by Thomas Jay Brown A thesis submitted in partial fulfillment of the requirements for the degree of Master of Arts in Anthropology Thesis Committee: Kenneth M. Ames, Chair Virginia L. Butler Shelby L. Anderson Portland State University 2016 © 2016 Thomas J. Brown i ABSTRACT Focusing on the relationship between demography and sedentary behavior, this thesis explores changes to mobility strategies on the Northern Northwest Coast of North America between 11,000 and 5,000 cal BP. Drawing on a regional database of radiocarbon dates, it uses summed probability distributions (SPDs) of calibrated dates as a proxy for population change, in combination with syntheses of previously published technological, paleo environmental and settlement pattern data to test three hypotheses derived from the literature about the development of logistic mobility among maritime hunter-gatherers on the Northern Coast. -

British Columbia Surficial Geology Map Index
60°N 138°W 136°W est. 1895 G E Y 134°W O E LO RV GICAL SU 114O 132°W 120°W 130°W 122°W 60°N 128°W 126°W 124°W British Columbia Geological Survey 114P Atlin Open File 2019-03 version 202001 104M Sheet 1 of 2 104N 104O 094P 59°N 104P 094M 094N 094O 114I 136°W 138°W British Columbia surficial 59°N 104L geology map index Fort Nelson 104K 104J Dease Lake 094I 104I 094L 094K 094J H. Arnold and T. Ferbey 134°W 58°N 58°N Surficial geology index by map scale Scale 1:1,000 to 1:49,999 Scale 1:200,000 to 1:299,999 104F Scale 1:50,000 to 1:99,999 Scale 1:300,000 to 1:1,000,000 104G 094H Scale 1:100,000 to 1:199,999 Scale 1:5,000,000 (entirety of province) 104H 094E 094F 094G Presented here is a surficial geology map index for British Columbia. These maps have been 57°N 57°N produced by the British Columbia Geological Survey, the Geological Survey of Canada (GSC), and 104C Geoscience BC. To be included in this index maps have to be available for digital download. Each map is represented by the actual map extent or footprint rather than a bounding box or NTS sheet that it falls within. This provides an accurate representation of the areal extent of surficial geology mapping for British Columbia. 104B 094A 104A 094D 094C 094B The index is presented by map scale. -

Heptacarpus Paludicola Class: Malacostraca Order: Decapoda a Broken Back Shrimp Section: Caridea Family: Thoridae
Phylum: Arthropoda, Crustacea Heptacarpus paludicola Class: Malacostraca Order: Decapoda A broken back shrimp Section: Caridea Family: Thoridae Taxonomy: Local Heptacarpus species (e.g. Antennae: Antennal scale never H. paludicola and H. sitchensis) were briefly much longer than rostrum. Antennular considered to be in the genus Spirontocaris peduncle bears spines on each of the three (Rathbun 1904; Schmitt 1921). However members of Spirontocaris have two or more segments and stylocerite (basal, lateral spine supraorbital spines (rather than only one in on antennule) does not extend beyond the Heptacarpus). Thus a known synonym for H. first segment (Wicksten 2011). paludicola is S. paludicola (Wicksten 2011). Mouthparts: The mouth of decapod crustaceans comprises six pairs of Description appendages including one pair of mandibles Size: Individuals 20 mm (males) to 32 mm (on either side of the mouth), two pairs of (females) in length (Wicksten 2011). maxillae and three pairs of maxillipeds. The Illustrated specimen was a 30 mm-long, maxillae and maxillipeds attach posterior to ovigerous female collected from the South the mouth and extend to cover the mandibles Slough of Coos Bay. (Ruppert et al. 2004). Third maxilliped without Color: Variable across individuals. Uniform expodite and with epipods (Fig. 1). Mandible with extremities clear and green stripes or with incisor process (Schmitt 1921). speckles. Color can be deep blue at night Carapace: No supraorbital spines (Bauer 1981). Adult color patterns arise from (Heptacarpus, Kuris et al. 2007; Wicksten chromatophores under the exoskeleton and 2011) and no lateral or dorsal spines. are related to animal age and sex (e.g. Rostrum: Well-developed, longer mature and breeding females have prominent than carapace, extending beyond antennular color patters) (Bauer 1981). -

OREGON ESTUARINE INVERTEBRATES an Illustrated Guide to the Common and Important Invertebrate Animals
OREGON ESTUARINE INVERTEBRATES An Illustrated Guide to the Common and Important Invertebrate Animals By Paul Rudy, Jr. Lynn Hay Rudy Oregon Institute of Marine Biology University of Oregon Charleston, Oregon 97420 Contract No. 79-111 Project Officer Jay F. Watson U.S. Fish and Wildlife Service 500 N.E. Multnomah Street Portland, Oregon 97232 Performed for National Coastal Ecosystems Team Office of Biological Services Fish and Wildlife Service U.S. Department of Interior Washington, D.C. 20240 Table of Contents Introduction CNIDARIA Hydrozoa Aequorea aequorea ................................................................ 6 Obelia longissima .................................................................. 8 Polyorchis penicillatus 10 Tubularia crocea ................................................................. 12 Anthozoa Anthopleura artemisia ................................. 14 Anthopleura elegantissima .................................................. 16 Haliplanella luciae .................................................................. 18 Nematostella vectensis ......................................................... 20 Metridium senile .................................................................... 22 NEMERTEA Amphiporus imparispinosus ................................................ 24 Carinoma mutabilis ................................................................ 26 Cerebratulus californiensis .................................................. 28 Lineus ruber ......................................................................... -

Hydrozoan Insights in Animal Development and Evolution Lucas Leclère, Richard Copley, Tsuyoshi Momose, Evelyn Houliston
Hydrozoan insights in animal development and evolution Lucas Leclère, Richard Copley, Tsuyoshi Momose, Evelyn Houliston To cite this version: Lucas Leclère, Richard Copley, Tsuyoshi Momose, Evelyn Houliston. Hydrozoan insights in animal development and evolution. Current Opinion in Genetics and Development, Elsevier, 2016, Devel- opmental mechanisms, patterning and evolution, 39, pp.157-167. 10.1016/j.gde.2016.07.006. hal- 01470553 HAL Id: hal-01470553 https://hal.sorbonne-universite.fr/hal-01470553 Submitted on 17 Feb 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Current Opinion in Genetics and Development 2016, 39:157–167 http://dx.doi.org/10.1016/j.gde.2016.07.006 Hydrozoan insights in animal development and evolution Lucas Leclère, Richard R. Copley, Tsuyoshi Momose and Evelyn Houliston Sorbonne Universités, UPMC Univ Paris 06, CNRS, Laboratoire de Biologie du Développement de Villefranche‐sur‐mer (LBDV), 181 chemin du Lazaret, 06230 Villefranche‐sur‐mer, France. Corresponding author: Leclère, Lucas (leclere@obs‐vlfr.fr). Abstract The fresh water polyp Hydra provides textbook experimental demonstration of positional information gradients and regeneration processes. Developmental biologists are thus familiar with Hydra, but may not appreciate that it is a relatively simple member of the Hydrozoa, a group of mostly marine cnidarians with complex and diverse life cycles, exhibiting extensive phenotypic plasticity and regenerative capabilities. -

1 Metagenetic Analysis of 2018 and 2019 Plankton Samples from Prince
Metagenetic Analysis of 2018 and 2019 Plankton Samples from Prince William Sound, Alaska. Report to Prince William Sound Regional Citizens’ Advisory Council (PWSRCAC) From Molecular Ecology Laboratory Moss Landing Marine Laboratory Dr. Jonathan Geller Melinda Wheelock Martin Guo Any opinions expressed in this PWSRCAC-commissioned report are not necessarily those of PWSRCAC. April 13, 2020 ABSTRACT This report describes the methods and findings of the metagenetic analysis of plankton samples from the waters of Prince William Sound (PWS), Alaska, taken in May of 2018 and 2019. The study was done to identify zooplankton, in particular the larvae of benthic non-indigenous species (NIS). Plankton samples, collected by the Prince William Sound Science Center (PWSSC), were analyzed by the Molecular Ecology Laboratory at the Moss Landing Marine Laboratories. The samples were taken from five stations in Port Valdez and nearby in PWS. DNA was extracted from bulk plankton and a portion of the mitochondrial Cytochrome c oxidase subunit 1 gene (the most commonly used DNA barcode for animals) was amplified by polymerase chain reaction (PCR). Products of PCR were sequenced using Illumina reagents and MiSeq instrument. In 2018, 257 operational taxonomic units (OTU; an approximation of biological species) were found and 60 were identified to species. In 2019, 523 OTU were found and 126 were identified to species. Most OTU had no reference sequence and therefore could not be identified. Most identified species were crustaceans and mollusks, and none were non-native. Certain species typical of fouling communities, such as Porifera (sponges) and Bryozoa (moss animals) were scarce. Larvae of many species in these phyla are poorly dispersing, such that they will be found in abundance only in close proximity to adult populations. -

National Historic Sites of Canada System Plan Will Provide Even Greater Opportunities for Canadians to Understand and Celebrate Our National Heritage
PROUDLY BRINGING YOU CANADA AT ITS BEST National Historic Sites of Canada S YSTEM P LAN Parks Parcs Canada Canada 2 6 5 Identification of images on the front cover photo montage: 1 1. Lower Fort Garry 4 2. Inuksuk 3. Portia White 3 4. John McCrae 5. Jeanne Mance 6. Old Town Lunenburg © Her Majesty the Queen in Right of Canada, (2000) ISBN: 0-662-29189-1 Cat: R64-234/2000E Cette publication est aussi disponible en français www.parkscanada.pch.gc.ca National Historic Sites of Canada S YSTEM P LAN Foreword Canadians take great pride in the people, places and events that shape our history and identify our country. We are inspired by the bravery of our soldiers at Normandy and moved by the words of John McCrae’s "In Flanders Fields." We are amazed at the vision of Louis-Joseph Papineau and Sir Wilfrid Laurier. We are enchanted by the paintings of Emily Carr and the writings of Lucy Maud Montgomery. We look back in awe at the wisdom of Sir John A. Macdonald and Sir George-Étienne Cartier. We are moved to tears of joy by the humour of Stephen Leacock and tears of gratitude for the courage of Tecumseh. We hold in high regard the determination of Emily Murphy and Rev. Josiah Henson to overcome obstacles which stood in the way of their dreams. We give thanks for the work of the Victorian Order of Nurses and those who organ- ized the Underground Railroad. We think of those who suffered and died at Grosse Île in the dream of reaching a new home.