Filamentous Algae Monitoring Program: Cacapon River Microcosm Study 2015
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morgan County Relocation Package
Morgan County Relocation Package Long & Foster/Webber & Associates, Realtors® 480 W. Jubal Early Drive, Suite 100 Winchester, Virginia 22601 Office: 540-662-3484 - Toll Free: 800-468-6619 www.webberrealty.com TABLE OF CONTENTS INTRODUCTION ---------------------------------------------------------------------------------2 GOVERNMENT -----------------------------------------------------------------------------------3 TAXES ---------------------------------------------------------------------------------------------4-5 LICENSE ------------------------------------------------------------------------------------------5-6 IMPORTANT NUMBERS -----------------------------------------------------------------------7 HEALTH ------------------------------------------------------------------------------------------8-9 CLIMATE ------------------------------------------------------------------------------------------10 POPULATION --------------------------------------------------------------------------------10-11 CHURCHES ---------------------------------------------------------------------------------------12 TEMPORARY LODGING -----------------------------------------------------------------12-14 SHOPPING ----------------------------------------------------------------------------------------15 TRANSPORTATION ---------------------------------------------------------------------------16 SCHOOLS -------------------------------------------------------------------------------------17-18 LIBRARIES ---------------------------------------------------------------------------------------19 -

02070001 South Branch Potomac 01605500 South Branch Potomac River at Franklin, WV 01606000 N F South Br Potomac R at Cabins, WV 01606500 So
Appendix D Active Stream Flow Gauging Stations In West Virginia Active Stream Flow Gauging Stations In West Virginia 02070001 South Branch Potomac 01605500 South Branch Potomac River At Franklin, WV 01606000 N F South Br Potomac R At Cabins, WV 01606500 So. Branch Potomac River Nr Petersburg, WV 01606900 South Mill Creek Near Mozer, WV 01607300 Brushy Fork Near Sugar Grove, WV 01607500 So Fk So Br Potomac R At Brandywine, WV 01608000 So Fk South Branch Potomac R Nr Moorefield, WV 01608070 South Branch Potomac River Near Moorefield, WV 01608500 South Branch Potomac River Near Springfield, WV 02070002 North Branch Potomac 01595200 Stony River Near Mount Storm,WV 01595800 North Branch Potomac River At Barnum, WV 01598500 North Branch Potomac River At Luke, Md 01600000 North Branch Potomac River At Pinto, Md 01604500 Patterson Creek Near Headsville, WV 01605002 Painter Run Near Fort Ashby, WV 02070003 Cacapon-Town 01610400 Waites Run Near Wardensville, WV 01611500 Cacapon River Near Great Cacapon, WV 02070004 Conococheague-Opequon 01613020 Unnamed Trib To Warm Spr Run Nr Berkeley Spr, WV 01614000 Back Creek Near Jones Springs, WV 01616500 Opequon Creek Near Martinsburg, WV 02070007 Shenandoah 01636500 Shenandoah River At Millville, WV 05020001 Tygart Valley 03050000 Tygart Valley River Near Dailey, WV 03050500 Tygart Valley River Near Elkins, WV 03051000 Tygart Valley River At Belington, WV 03052000 Middle Fork River At Audra, WV 03052450 Buckhannon R At Buckhannon, WV 03052500 Sand Run Near Buckhannon, WV 03053500 Buckhannon River At Hall, WV 03054500 Tygart Valley River At Philippi, WV Page D 1 of D 5 Active Stream Flow Gauging Stations In West Virginia 03055500 Tygart Lake Nr Grafton, WV 03056000 Tygart Valley R At Tygart Dam Nr Grafton, WV 03056250 Three Fork Creek Nr Grafton, WV 03057000 Tygart Valley River At Colfax, WV 05020002 West Fork 03057300 West Fork River At Walkersville, WV 03057900 Stonewall Jackson Lake Near Weston, WV 03058000 West Fork R Bl Stonewall Jackson Dam Nr Weston 03058020 West Fork River At Weston, WV 03058500 W.F. -

The Cacapon Settlement: 1749-1800 31
THE CACAPON SETTLEMENT: 1749-1800 31 THE CACAPON SETTLEMENT: 1749-1800 31 5 THE CACAPON SETTLEMENT: 1749-1800 The existence of a settlement of Brethren families in the Cacapon River Valley of eastern Hampshire County in present day West Virginia has been unknown and uninvestigated until the present time. That a congregation of Brethren existed there in colonial times cannot now be denied, for sufficient evidence has been accumulated to reveal its presence at least by the 1760s and perhaps earlier. Because at this early date, Brethren churches and ministers did not keep records, details of this church cannot be recovered. At most, contemporary researchers can attempt to identify the families which have the highest probability of being of Brethren affiliation. Even this is difficult due to lack of time and resources. The research program for many of these families is incomplete, and this chapter is offered tentatively as a basis for additional research. Some attempted identifications will likely be incorrect. As work went forward on the Brethren settlements in the western and southern parts of old Hampshire County, it became clear that many families in the South Branch, Beaver Run and Pine churches had relatives who had lived in the Cacapon River Valley. Numerous families had moved from that valley to the western part of the county, and intermarriages were also evident. Land records revealed a large number of family names which were common on the South Branch, Patterson Creek, Beaver Run and Mill Creek areas. In many instances, the names appeared first on the Cacapon and later in the western part of the county. -

Program Overview
WWeett WWaaddeerrss aanndd BBeeyyoonndd TThhee CCoonnddiittiioonn ooff OOuurr SSttaattee’’ss WWaatteerrss AA CCiittiizzeenn’’ss PPeerrssppeeccttiivvee 1 WV Department of Environmental Protection Division of Water and Waste Management, Nonpoint Section 601 57th Street, SE Charleston, WV 25304 The document was prepared by Tim Craddock, WV DEP’s Citizens’ Monitoring Coordinator and is available electronically in Portable Document Format (PDF). To request your copy send e-mail to Tim Craddock at: [email protected]. ACKNOWLEDGEMENTS Color photographs provided by: Alana Hartman, DEP’s Potomac Basin Coordinator; Abby Chappel, WV River Network; Sherry Evasic, Blue Heron Environmental Network; Neil Gillies, Cacapon Institute; Suzanne Hubbard, The Mountain Institute; Renee Cain, Lower West Fork Watershed Association; Martin Christ, Friends of Deckers Creek; Bobby Bonnett, Heizer-Manila Watershed Organization; Diana Green, Davis Creek Watershed Association; James Grey, Morris Creek Watershed Association; Larry Orr, Kanawha Valley Chapter of Trout Unlimited; Valerie Wilson, Science Teacher, Oak Hill Catholic Center; Brad Durst, WV Conservation Agency and Curtis Canada, Upper Guyandotte Watershed Association. WV Save Our Streams would like to recognize all the volunteer monitors, not only those directly associated with the program, but any others who have given their time and energy in an effort to protect our state’s streams and rivers. WV Save Our Streams would also like to recognize all of the agency and other partners who have provided assistance of any kind, to help guide volunteers through the myriad of processes involved with water quality issues. “Perception is not acquired by formal education, nor is it reserved for persons learned in the arts or sciences. -

Spring 2021 Issue
West Virginia's Chesapeake Bay Trib Team Spring 2021 | Issue 39 Highlights and News Latest updates and stories related to WV progress. WV Highlights Discover: Jennings Randolph Lake Subscribe to our newsletter! Jennings Randolph Lake, located on the North Branch of the Potomac River between Maryland Sign up now! and West Virginia, provides water quality improvements, reduces flood damages downstream, provides a source of water supply for Find us on Facebook! downstream municipalities and industries, and provides the public with recreation opportunities. READ MORE New Employee Spotlight Shared from: WV WaterNet: Spring Edition Be sure to sign up for their e- newsletter! WVDEP’s Save Our Streams Program Nets New Coordinator Seeks Volunteers Ready to Monitor Stream Health through Benthic Macroinvertebrate Surveys The West Virginia Department of Environmental Protection’s (WVDEP) Watershed Improvement Branch welcomed Callie Cronin Sams as the new Save Our Streams (SOS) program coordinator in February. Read More Read More Our Partners are Doing Great Work! Rain Station Shepherdstown partnered with the Eastern Panhandle Planning and Development Council (Region 9) to install green infrastructure practices at the former train station campus. READ MORE Cacapon Watershed Project A dedicated source of funding has been directed to environmental practices in the Cacapon & Lost Rivers watershed, thanks to a successful effort by the USDA Natural Resources Conservation Service (NRCS) and local stakeholders. The project is known as a Focused Conservation Approach, which means that a portion of NRCS Environmental Quality Incentives Program (EQIP) funding, about $600,000 over 3 years, will be directed to this specific watershed. Text Link READ MORE WV DEP Gathering Information About Potential Pollutant Sources in Cacapon River Watershed Scientists from the WV Department of Environmental Protection's Watershed Assessment Branch plan to monitor select streams in the Cacapon River Watershed beginning in June 2021 and extending for approximately one year. -

Fishing Regulations JANUARY - DECEMBER 2004
WEST VIRGINIA Fishing Regulations JANUARY - DECEMBER 2004 West Virginia Division of Natural Resources D I Investment in a Legacy --------------------------- S West Virginia’s anglers enjoy a rich sportfishing legacy and conservation ethic that is maintained T through their commitment to our state’s fishery resources. Recognizing this commitment, the R Division of Natural Resources endeavors to provide a variety of quality fishing opportunities to meet I increasing demands, while also conserving and protecting the state’s valuable aquatic resources. One way that DNR fulfills this part of its mission is through its fish hatchery programs. Many anglers are C aware of the successful trout stocking program and the seven coldwater hatcheries that support this T important fishery in West Virginia. The warmwater hatchery program, although a little less well known, is still very significant to West Virginia anglers. O West Virginia’s warmwater hatchery program has been instrumental in providing fishing opportunities F to anglers for more than 60 years. For most of that time, the Palestine State Fish Hatchery was the state’s primary facility dedicated to the production of warmwater fish. Millions of walleye, muskellunge, channel catfish, hybrid striped bass, saugeye, tiger musky, and largemouth F and smallmouth bass have been raised over the years at Palestine and stocked into streams, rivers, and lakes across the state. I A recent addition to the DNR’s warmwater hatchery program is the Apple Grove State Fish Hatchery in Mason County. Construction of the C hatchery was completed in 2003. It was a joint project of the U.S. Army Corps of Engineers and the DNR as part of a mitigation agreement E for the modernization of the Robert C. -
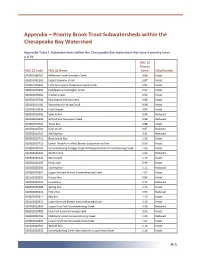
Appendix – Priority Brook Trout Subwatersheds Within the Chesapeake Bay Watershed
Appendix – Priority Brook Trout Subwatersheds within the Chesapeake Bay Watershed Appendix Table I. Subwatersheds within the Chesapeake Bay watershed that have a priority score ≥ 0.79. HUC 12 Priority HUC 12 Code HUC 12 Name Score Classification 020501060202 Millstone Creek-Schrader Creek 0.86 Intact 020501061302 Upper Bowman Creek 0.87 Intact 020501070401 Little Nescopeck Creek-Nescopeck Creek 0.83 Intact 020501070501 Headwaters Huntington Creek 0.97 Intact 020501070502 Kitchen Creek 0.92 Intact 020501070701 East Branch Fishing Creek 0.86 Intact 020501070702 West Branch Fishing Creek 0.98 Intact 020502010504 Cold Stream 0.89 Intact 020502010505 Sixmile Run 0.94 Reduced 020502010602 Gifford Run-Mosquito Creek 0.88 Reduced 020502010702 Trout Run 0.88 Intact 020502010704 Deer Creek 0.87 Reduced 020502010710 Sterling Run 0.91 Reduced 020502010711 Birch Island Run 1.24 Intact 020502010712 Lower Three Runs-West Branch Susquehanna River 0.99 Intact 020502020102 Sinnemahoning Portage Creek-Driftwood Branch Sinnemahoning Creek 1.03 Intact 020502020203 North Creek 1.06 Reduced 020502020204 West Creek 1.19 Intact 020502020205 Hunts Run 0.99 Intact 020502020206 Sterling Run 1.15 Reduced 020502020301 Upper Bennett Branch Sinnemahoning Creek 1.07 Intact 020502020302 Kersey Run 0.84 Intact 020502020303 Laurel Run 0.93 Reduced 020502020306 Spring Run 1.13 Intact 020502020310 Hicks Run 0.94 Reduced 020502020311 Mix Run 1.19 Intact 020502020312 Lower Bennett Branch Sinnemahoning Creek 1.13 Intact 020502020403 Upper First Fork Sinnemahoning Creek 0.96 -

Center for Excellence in Disabilities at West Virginia University, Robert C
ACKNOWLEDGEMENTS This publication was made possible by the support of the following organizations and individuals: Center for Excellence in Disabilities at West Virginia University, Robert C. Byrd Health Sciences Center West Virginia Assistive Technology System (WVATS) West Virginia Division of Natural Resources West Virginia Division of Tourism Partnerships in Assistive Technologies, Inc. (PATHS) Special thanks to Stephen K. Hardesty and Brittany Valdez for their enthusiasm while working on this Guide. 1 Table of Contents INTRODUCTION .......................................................... 3 • How to Use This Guide ......................................... 4 • ADA Sites .............................................................. 5 • Types of Fish ......................................................... 7 • Traveling in West Virginia ...................................... 15 COUNTY INDEX .......................................................... 19 ACTIVITY LISTS • Public Access Sites ............................................... 43 • Lakes ..................................................................... 53 • Trout Fishing ......................................................... 61 • River Float Trips .................................................... 69 SITE INDEX ................................................................. 75 SITE DESCRIPTIONS .................................................. 83 APPENDICES A. Recreation Organizations ......................................207 B. Trout Stocking Schedule .......................................209 -

Development of Outdoor Recreation Resource Amenity Indices for West Virginia
Graduate Theses, Dissertations, and Problem Reports 2008 Development of outdoor recreation resource amenity indices for West Virginia Jing Wang West Virginia University Follow this and additional works at: https://researchrepository.wvu.edu/etd Recommended Citation Wang, Jing, "Development of outdoor recreation resource amenity indices for West Virginia" (2008). Graduate Theses, Dissertations, and Problem Reports. 2680. https://researchrepository.wvu.edu/etd/2680 This Thesis is protected by copyright and/or related rights. It has been brought to you by the The Research Repository @ WVU with permission from the rights-holder(s). You are free to use this Thesis in any way that is permitted by the copyright and related rights legislation that applies to your use. For other uses you must obtain permission from the rights-holder(s) directly, unless additional rights are indicated by a Creative Commons license in the record and/ or on the work itself. This Thesis has been accepted for inclusion in WVU Graduate Theses, Dissertations, and Problem Reports collection by an authorized administrator of The Research Repository @ WVU. For more information, please contact [email protected]. Development of Outdoor Recreation Resource Amenity Indices for West Virginia Jing Wang Thesis submitted to the Davis College of Agriculture, Forestry, and Consumer Sciences At West Virginia University in partial fulfillment of the requirements for the degree of Master of Science in Recreation, Parks, and Tourism Resources Jinyang Deng, Ph.D., Chair Chad -

Regulations Summary 2019
WEST VIRGINIA FISHINGRegulations Summary 2019 wvdnr.gov From the Director Last year the DNR released an updated, online interactive map that provides valuable information on all aspects of fishing and hunting adventures. DNR personnel are continuing to update information and produce new, useful maps. After hearing about the need from anglers in an online survey, DNR personnel collected lake depth data and processed new bathymetry maps for 35 lakes across the state. These maps are now available on the interactive fishing map and downloadable to print or take with you on your mobile device. Also, anglers can now access the real-time streamflow conditions from the U.S. Geological Survey on our interactive fishing map. The real-time information allows anglers to check on flow and make decisions about whether fishing conditions are ideal before heading out to a stream or river. Visit wvdnr.gov/gis for more details and links to the interactive map and other map pages. You helped fund this project through the Sport Fish Restoration Program, using excise taxes on selected fishing equipment and boat fuel. I encourage you to take advantage of West Virginia’s abundant natural resources and go fishing every chance you get. And take a friend or family member with you. It’s a great way to relax and enjoy each other’s company. Stephen S. McDaniel, DNR Director DISTRICT OFFICES Main Office - South Charleston, WV 25303 324 4th Avenue Fish Management − Mark T. Scott (304) 558-2771 Law Enforcement − Col. Jerry Jenkins (304) 558-2784 License Unit − Michael Ingram (304) 558-2758 District 1 - Farmington, WV 26571 1110 Railroad Street (304) 825-6787 Fish Management − Dave Wellman Law Enforcement − Capt. -

Conserving Habitat Through the FEDERAL Farm Bill a Guide for Land Trusts and Landowners Acknowledgements
conserving habitat through the FEDERAL farm bill A Guide for Land Trusts and Landowners Acknowledgements principal author Aimee Weldon Living Lands Project Manager, Defenders of Wildlife (2007-2009) Senior Director of Restoration and Lands, Potomac Conservancy (2009-Present) contributing authors Randall Gray, Farm Bill Coordinator, Intermountain West Joint Venture Ferd Hoefner, Policy Director, National Sustainable Agriculture Coalition Brad Redlin, Agricultural Program Director, Izaak Walton League of America Judy Boshoven, Living Lands Project Manager, Defenders of Wildlife The authors gratefully acknowledge the generous financial support of the Biophilia Foundation and the Living Lands Project at Defenders of Wildlife. For contributions of technical content, we thank the Intermountain West Joint Venture and the North American Bird Conservation Initiative. We also thank the many land trusts, landowners and other partners who so generously provided the case studies and personal experiences that bring this guide to life. And to the millions of dedicated and thoughtful farmers, ranchers, foresters and partners who are committed to caring for the land and all of its inhabitants, we are deeply indebted. The following partners contributed to the content or production of this guide: Defenders of Wildlife | Intermountain West Joint Venture | Izaak Walton League of America National Sustainable Agriculture Coalition | North American Bird Conservation Initiative | Potomac Conservancy We also thank the following individuals for their assistance with this guide: Aimee Delach, Amielle DeWan, Noah Kahn, Sara O’Brien, Pine Roehrs, Allison Barra Srinivas Editorial Staff: Kate Davies, Editor; Charles Kogod, Photo Editor; Peter Corcoran, Designer For additional information, contact: Aimee Weldon Potomac Conservancy 8601 Georgia Avenue Suite 612 Silver Spring, Maryland 20910 301-608-1188, ext. -

Top10 Highest Historical Crests: Cacapon River Near Great Cacapon, WV
Top10 Highest Historical Crests: Cacapon River near Great Cacapon, WV Latitude: 39.582 Period of Record: 1889-Present Longitude: -78.31 Flood Stage: 9 Last Flood: 12/22/2018 Number of Floods: 93 Date of Flood Crest (ft) Streamflow (cfs) Weather Summary 3/18/1936 30.1 87,600 Two successive rainstorms combined with snowmelt flooded the Eastern Seaboard from Virginia to Maine. A total of 150 to 200 lives were lost and damage estimates were over $1 million. 6/1/1889 24.7 57,500 A mesoscale convective complex caused 9 inches of rain to fall in 36 hours with 4 inches of rain occurring in 8 hours. Most of Williamsport, Lock Haven, Muncy, Jersey Shore and Montgomery were submerged. 8/19/1955 24.3 55,500 Hurricane Diane made landfall 5 days after Hurricane Connie. Hurricane Diane produced several inches of rain with locally heavier amounts of 10 to 20 inches. 10/16/1942 23.67 52,600 The remnants of the eighth tropical storm of the year produced torrential rains and caused the worst river flooding in the history of Virginia. 4/26/1937 22.6 47,400 Heavy rains caused widespread flooding across most of Virginia. The Rappahannock basin was hit the hardest, but the Potomac River at Wisconsin Avenue rose to 14.3 feet. Six people died and total damages were well over $1.5 million. 6/22/1972 22.17 45,500 Hurricane Agnes made landfall again over southeastern New York on June 22 and moved westward into Pennsylvania. Rainfall totals from June 20-25 range from 2-3 inches in the Upper Potomac to 18 inches near Shamokin, Pennsylvania.