I Marshelder (Iva Annua
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Noxious and Rangeland Weed Management: Getting the Most out of Spraying Mark Pederson Dow Agrosciences [email protected]
Noxious and Rangeland Weed Management: Getting the Most Out of Spraying Mark Pederson Dow AgroSciences [email protected] DOW RESTRICTED - For internal use only Thanks to Pend Oreille County. DOW RESTRICTED - For internal use only Successful Herbicide Weed Control • Why does the herbicide work better one year and not the next? • What changes from year to year? • Is your equipment in good working order? • Just keeping on doing what you always have done… • And get what you always got! DOW RESTRICTED - For internal use only The Variables • Weather • Growing conditions • Temperatures • Wind • Water volume per acre applied • Equipment wear and calibration • You, you, you DOW RESTRICTED - For internal use only Start with the End in Mind • Visualize what you want to accomplish with every spray job – set expectations • Review records and see what has changed • Are the growing conditions and weeds conducive to weed control? • Recalibrate every year and even do a check up during the spray season • Calibrate every applicator that applies products DOW RESTRICTED - For internal use only Rates, Carrier & Equipment • Select herbicide product that: – controls the majority of target weeds – use the correct rates (don’t scrimp) • Water carrier per acre – use enough to get the product to the desired site of activity (if herbicide is soil active, it must penetrate the vegetative canopy to get benefit) – Use Syltac or R-11 • Tune-up sprayers and do timely repairs DOW RESTRICTED - For internal use only Adjuvants/Surfactants • Always use Syltac or R-11 • Use -

The Future of the Past: Science in Archaeology Illinois Antiquity Vol
The Future of the Past: Science in Archaeology Illinois Antiquity Vol. 50, No. 3 September 2015 REFERENCES AND SUGGESTED READINGS ARCHAEOLOGY AND ECOLOGY: BRIDGING THE SCIENCES THROUGH INTERDISCIPLINARY RESEARCH By Carol E. Colaninno LiDAR ILLUMINATED By Michael Farkas IDENTIFYING BLACK DRINK CEREMONIALISM AT CAHOKIA: CHEMICAL RESIDUE ANALYSIS By Thomas E. Emerson and Timothy R. Pauketat SOURCING NATIVE AMERICAN CERAMICS FROM WESTERN ILLINOIS By Julie Zimmermann Holt, Andrew J. Upton, and Steven A. Hanlin Conrad, Lawrence A. 1989 The Southeastern Ceremonial Complex on the Northern Middle Mississippian Frontier: Late Prehistoric Politico-religious Systems in the Central Illinois River Valley. In The Southeastern Ceremonial Complex: Artifacts and Analysis, edited by P. Galloway, pp. 93-113. University of Nebraska Press, Lincoln. 1991 The Middle Mississippian Cultures of the Central Illinois Valley. In Cahokia and the Hinterlands: Middle Mississippian Cultures of the Midwest, edited by T. E. Emerson and R. B. Lewis, pp. 119-156. University of Illinois Press, Urbana. Dye, David H. 2004 Art, Ritual, and Chiefly Warfare in the Mississippian World. In Hero, Hawk, and Open Hand: American Indian Art of the Ancient Midwest and South, edited by R. F. Townsend, pp. 191-205. The Art Institute, Chicago. Fie, Shannon M. 2006 Visiting in the Interaction Sphere: Ceramic Exchange and Interaction in the Lower Illinois Valley. In Recreating Hopewell, edited by D. K. Charles and J. E. Buikstra, pp. 427-45. University Press of Florida, Gainesville. 2008 Middle Woodland Ceramic Exchange in the Lower Illinois Valley. Midcontinental Journal of Archaeology 33:5-40. Fowles, Severin M., Leah Minc, Samuel Duwe and David V. -

Crops Before Corn an INVESTIGATION of EASTERN AGRICULTURAL COMPLEX PLANTS at the TOM JONES SITE, ARKANSAS
Crops Before Corn AN INVESTIGATION OF EASTERN AGRICULTURAL COMPLEX PLANTS AT THE TOM JONES SITE, ARKANSAS ROSIE BLEWITT-GOLSCH Corn, Beans, and Squash Eastern Agricultural Complex • Initial domestication ca. 4000 – 3000 BP • Earliest occurrences found in upper South and lower Midwest • Initial evidence of EAC from rock shelters, but difficult to prove that they were actually grown as crops in prehistory Wild-Cultivated-Domesticated Spectrum Activities Outcomes • No intervention • Cultivation - preparing a seed bed, garden, field, • Encourage/tend planting the seeds Clearing out competing however, identical trees to wild plants Bringing water to plants • Domestication - putting during drought different selective Pruning plants pressures on plants and animals; generally genetic Burning and/or phenotypic • Control reproduction plants dependent Transplanting on people Collecting and planting seeds Eastern Agricultural Complex Plant Species • Sumpweed (Iva annua) • Sunflower (Helianthus annus) • Maygrass (Phalaris caroliniana) • Goosefoot (Chenopodium berlandieri) • Knotweed (Polygonum erectum) • Little Barley (Hordeum pusillum) • Squash (Cucurbita pepo) Squash and Gourds • Bottle gourd (date?) • Squash (4400 BP) • Produce starchy seeds but not much flesh • Rinds can be used as floats or containers • Domestication marked by seed enlargement and increase in rind thickness Squash Bottle Gourd Sunflower and Sumpweed • Sunflower (4300 BP) • Sumpweed (3900 BP) • Oily seeds • Domestication marked by increase in seed size Sunflower Sumpweed Goosefoot and Erect Knotweed • Goosefoot (3500 BP) • Erect knotweed (2500 BP) • Starchy seeds, Fall food source • Seed coat thickness, larger perisperm Knotweed Goosefoot Goosefoot Wild goosefoot Domesticated goosefoot Domesticated? variant Wild? variant Erect knotweed – two phenotypes Longer/shorter Thick/thin seed coat Maygrass and Little Barley • Maygrass (date?) • Little barley (date?) • Starchy seeds, Spring resources • Geographic range, quantity little barley Maygrass WHEAT Middle East Domesticated ~7,000 B.C. -

CHROMOSOME NUMBERS in COMPOSITAE. IV. Ambrosieael
Amer, Jour. Bot. 51(4): 419-424.1964. CHROMOSOME NUMBERS IN COMPOSITAE. IV. AMBROSIEAEl WILLARD W. PAYNE, PETER H. RAVEN AND DONALD W. KYHOS Department of Botany, The University of Michigan, Ann Arbor, Michigan and Division of Systematic Biology, Stanford University, Stanford, California ABSTRACT New chromosome observations are reported for 30 species and varieties from 4 genera of Am brosieae: Ambrosia (including Franseria), Dicoria, Humenoclea and Iva. Neither polyploidy nor aneuploidy is known in the genera Dicorea, Hymenoclea or Xanthium. Aneuplcid reduction appears to have played a role in the genome evolution of several species of Iva and Ambrosia. Polyploid species occur in both Iva and Ambrosia and polyploid series exist for at least 5 species or species aggregates of the latter. All available evidence indicates that the primitive chromosome number for the tribe is x = 18, differentiation and speciation having occurred at this level, which is here termed diploid. The group, however, must ultimately have been of polyploid origin from forms with z = 9. THIS is the fourth in a recent series of papers (Payne, 1963). For the present, therefore, we have dealing with chromosome numbers in various deemed it the best course to recognize the group tribes of the Compositae. Materials and methods as a distinct tribe, the Ambrosieae, a treatment are generally those discussed in earlier contribu which follows the precedents of Cassini (1834) and tions to this series, the most recent of which is Delpino (1871), and which agrees in general Ornduff et al. (1963). Procedural details have meaning with the remarks or treatments of varied somewhat since work was carried out Bentham (1873), Small (1913), Rydberg (1922) independently by the various workers. -

Old Stone Fort Archaeological Society March 2015
Old Stone Fort Archaeological Society March 2015 The Old Stone Fort Archaeological Society will meet Thursday, March 12th at 6:00 pm at the Old Stone Fort Museum in Manchester, Tennessee. The purpose of this Society is to learn more about prehistoric Native American cultures and the early historic period of Tennessee. Our meetings are scheduled for the second Thursday of each month and are free and open to the public. Our program this month: Mark Norton who will present Big Bone Cave Big Bone Cave is located in Van Buren County, Tennessee and has a long history of habitation/exploitation by ice age animals, Native Americans, and saltpeter miners. Extinct Ground Sloth and Jaguar remains have been recovered from this large cave as well as a number of ancient birds. Cavers and archaeologists have also Big Bone Cave recovered numerous cane torches, moccasins, and other Managed by Rock Island State Park artifacts of prehistoric Native Americans. The historic http://tnstateparks.com/parks/about/rock-island period exploitation of this cave was in the quest to obtain saltpeter, first during the War of 1812 and later during the Civil War. This presentation will highlight each of these archaeologically significant periods discovered within Big Bone Cave. This cave is currently closed due to the white nose syndrome that has infected the native bat populations of our region. Rock Island State Park manages this natural area and hopes to reopen this fascinating cave for tours once the white nose problem has been solved. See you Thursday, Mark Information: -
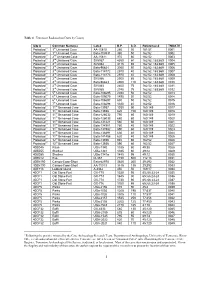
Table 4. Tennessee Radiocarbon Dates by County Site # Common
Table 4. Tennessee Radiocarbon Dates by County Site # Common Name(s) Lab # B.P. S.D. Reference # TNDATE Protected 1 1st Unnamed Cave AA-15810 260 50 161:57 0001 Protected 1 1st Unnamed Cave Beta-103531 380 50 162:52 0002 Protected 1 2nd Unnamed Cave AA-15811 970 60 162:52 0003 Protected 1 3rd Unnamed Cave SI-5067 4350 60 162:52; 163:669 0004 Protected 1 3rd Unnamed Cave SI-5064 3115 65 162:52; 163:669 0005 Protected 1 3rd Unnamed Cave Beta-96624 3060 50 162:52; 163:669 0006 Protected 1 3rd Unnamed Cave Beta-114172 2970 40 162:52; 163:669 0007 Protected 1 3rd Unnamed Cave Beta-114173 2970 40 162:52; 163:669 0008 Protected 1 3rd Unnamed Cave SI-5066 2950 65 162:52; 163:669 0009 Protected 1 3rd Unnamed Cave Beta-96623 2950 110 162:52; 163:669 0010 Protected 1 3rd Unnamed Cave SI-5063 2805 75 162:52; 163:669 0011 Protected 1 3rd Unnamed Cave SI-5065 2745 75 162:52; 163:669 0012 Protected 1 5th Unnamed Cave Beta-106695 2030 50 162:52 0013 Protected 1 6th Unnamed Cave Beta-109675 1890 50 162:52 0014 Protected 1 6th Unnamed Cave Beta-106697 630 50 162:52 0015 Protected 1 7th Unnamed Cave Beta-106698 1320 40 162:52 0016 Protected 1 11th Unnamed Cave Beta-13937 1030 90 160:149 0017 Protected 1 11th Unnamed Cave Beta-13938 620 150 160:149 0018 Protected 1 11th Unnamed Cave Beta-126032 750 60 160:149 0019 Protected 1 11th Unnamed Cave Beta-126033 680 60 160:149 0020 Protected 1 11th Unnamed Cave Beta-131221 560 60 160:149 0021 Protected 1 11th Unnamed Cave Beta 134981 780 40 160:149 0022 Protected 1 11th Unnamed Cave Beta-134982 890 60 160:149 0023 -

A Northern Nevada Homeowner's Guide to Identifying and Managing
Fact Sheet-13-15 A Northern Nevada Homeowner’s Guide to Identifying and Managing Poverty Sumpweed Susan Donaldson, Water Quality and Weed Specialist Wendy Hanson Mazet, Master Gardener Program Coordinator and Horticulturist Other common names: Povertyweed, bozzleweed, death- weed, salt sage, small-flowered marsh elder Scientific name: Iva axillaris Family: Asteraceae Description: Poverty sumpweed grows to 2 feet tall in large colonies in disturbed sites. The leaves have a strong odor most people find unpleasant. When touched, the foliage can cause contact dermatitis in sensitive people. Typical plant growing in a Leaves: Grayish-green, lobed and covered with short, bristly disturbed site. hairs. Oppositely attached to the stem toward the bottom of the plant, and alternately attached above. Stems: Gray-green, bristly, upright and branched. Flowers: Produces greenish male and female flowers on the same plant during the summer. Male flowers hang or nod at the ends of branches. Female flowers are spiny and are found in the leaf axils. Roots: Grows deep, woody creeping roots. Native to: Western North America Where it grows: Salt marshes, alkali plains, roadsides and pastures, and sites disturbed by cultivation or overgrazing Plants can sprout from dormant roots Life cycle: Perennial (lives longer than two years) after long periods. Seedlings are uncommon. Reproduction: Reproduces by creeping roots and seed (Top photo by S. Donaldson; bottom photo by W. Hanson Mazet) Control methods: Poverty sumpweed can be difficult to control due to its deep, spreading root system. As with all perennials, the roots must be killed to effectively control this plant. Mechanical: Cultivation may spread root. -

Archaeological Survey of the Chesapeake Bay Shorelines Associated with Accomack County and Northampton County, Virginia
ARCHAEOLOGICAL SURVEY OF THE ATLANTIC COAST SHORELINES ASSOCIATED WITH ACCOMACK COUNTY AND NORTHAMPTON COUNTY, VIRGINIA Survey and Planning Report Series No. 7 Virginia Department of Historic Resources 2801 Kensington Avenue Richmond, VA 23221 2003 ARCHAEOLOGICAL SURVEY OF THE ATLANTIC COAST SHORELINES ASSOCIATED WITH ACCOMACK COUNTY AND NORTHAMPTON COUNTY, VIRGINIA Virginia Department of Historic Resources Survey and Planning Report Series No. 7 Author: Darrin L. Lowery Chesapeake Bay Watershed Archaeological Research Foundation 5264 Blackwalnut Point Road, P.O. Box 180 Tilghman, MD 21671 2003 ii ABSTRACT This report summarizes the results of an archaeological survey conducted along the Atlantic shorelines of both Accomack County and Northampton County, Virginia. Accomack and Northampton Counties represent the southernmost extension of the Delmarva Peninsula. The study area encompasses all of the lands adjacent to the Atlantic Ocean and shorelines associated with the back barrier island bays. A shoreline survey was conducted along the Atlantic Ocean to gauge the erosion threat to the archaeological resources situated along the shoreline. Archaeological sites along shorelines are subjected to numerous natural processes which hinder site visibility and limit archaeological interpretations. Summaries of these natural processes are presented in this report. The primary goal of the project was to locate, identify, and record any archaeological sites or remains along the Atlantic seashore that are threatened by shoreline erosion. The project also served as a test of a prehistoric site predictive/settlement model that has been utilized during other archaeological surveys along the Chesapeake Bay shorelines and within the interior sections of the Delmarva Peninsula. The prehistoric site predictive/settlement model is presented in detail using archaeological examples from Maryland and Virginia’s Eastern Shore. -

Bone Flutes and Whistles from Archaeological Sites in Eastern North America
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Masters Theses Graduate School 12-1976 Bone Flutes and Whistles from Archaeological Sites in Eastern North America Katherine Lee Hall Martin University of Tennessee - Knoxville Follow this and additional works at: https://trace.tennessee.edu/utk_gradthes Part of the Anthropology Commons Recommended Citation Martin, Katherine Lee Hall, "Bone Flutes and Whistles from Archaeological Sites in Eastern North America. " Master's Thesis, University of Tennessee, 1976. https://trace.tennessee.edu/utk_gradthes/1226 This Thesis is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Masters Theses by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a thesis written by Katherine Lee Hall Martin entitled "Bone Flutes and Whistles from Archaeological Sites in Eastern North America." I have examined the final electronic copy of this thesis for form and content and recommend that it be accepted in partial fulfillment of the equirr ements for the degree of Master of Arts, with a major in Anthropology. Charles H. Faulkner, Major Professor We have read this thesis and recommend its acceptance: Major C. R. McCollough, Paul W . Parmalee Accepted for the Council: Carolyn R. Hodges Vice Provost and Dean of the Graduate School (Original signatures are on file with official studentecor r ds.) To the Graduate Council: I am submitting herewith a thesis written by Katherine Lee Hall Mar tin entitled "Bone Flutes and Wh istles from Archaeological Sites in Eastern North America." I recormnend that it be accepted in partial fulfillment of the requirements for the degree of Master of Arts, with a maj or in Anthropology. -

Discover Illinois Archaeology
Discover Illinois Archaeology ILLINOIS ASSOCIATION FOR ADVANCEMENT OF ARCHAEOLOGY ILLINOIS ARCHAEOLOGICAL SURVEY Discover Illinois Archaeology Illinois’ rich cultural heritage began more collaborative effort by 18 archaeologists from than 12,000 years ago with the arrival of the across the state, with a major contribution by ancestors of today’s Native Americans. We learn Design Editor Kelvin Sampson. Along with sum- about them through investigations of the remains maries of each cultural period and highlights of they left behind, which range from monumental regional archaeological research, we include a earthworks with large river-valley settlements to short list of internet and print resources. A more a fragment of an ancient stone tool. After the extensive reading list can be found at the Illinois arrival of European explorers in the late 1600s, a Association for Advancement of Archaeology succession of diverse settlers added to our cul- web site www.museum.state.il.us/iaaa/DIA.pdf. tural heritage, leading to our modern urban com- We hope that by reading this summary of munities and the landscape we see today. Ar- Illinois archaeology, visiting a nearby archaeo- chaeological studies allow us to reconstruct past logical site or museum exhibit, and participating environments and ways of life, study the rela- in Illinois Archaeology Awareness Month pro- tionship between people of various cultures, and grams each September, you will become actively investigate how and why cultures rise and fall. engaged in Illinois’ diverse past and DISCOVER DISCOVER ILLINOIS ARCHAEOLOGY, ILLINOIS ARCHAEOLOGY. summarizing Illinois culture history, is truly a Alice Berkson Michael D. Wiant IIILLINOIS AAASSOCIATION FOR CONTENTS AAADVANCEMENT OF INTRODUCTION. -

A Comparative Study of Oneota and Langford Traditions
A COMPARATIVE STUDY OF ONEOTA AND LANGFORD TRADITIONS by Chrisie L. Hunter A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science in Anthropology at The University of Wisconsin-Milwaukee May 2002 A COMPARATIVE STUDY OF ONEOTA AND LANGFORD TRADITIONS by Chrisie L. Hunter A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science in Anthropology at The University of Wisconsin-Milwaukee May 2002 ________________________________________________________________________ Major Professor Date ________________________________________________________________________ Graduate School Approval Date ii ABSTRACT A COMPARATIVE STUDY OF ONEOTA AND LANGFORD TRADITIONS by Chrisie L. Hunter The University of Wisconsin-Milwaukee, 2002 Under the Supervision of Dr. Robert J. Jeske This study is a comparative analysis of two contemporaneous sites from two related cultural systems, Oneota and Langford. Similarities and differences between the sites were examined through analysis of technology, faunal and floral remains, and environmental variables. A catchment analysis was completed to understand environmental factors affecting agricultural practices and resource utilization between the two sites. The issue of agricultural production and the extent this subsistence strategy was utilized at two sites was examined. The Crescent Bay Hunt Club site (Oneota) and Washington Irving site (Langford) are shown to be similar in lithic technology but significantly different in ceramic technology, and subsistence strategies. The Oneota Crescent Bay Hunt Club site occupants were more dependant upon wetland resources than were the Langford occupants of the Washington Irving site. _______________________________________________________________________ Major Professor Date iii © Copyright by Chrisie L. Hunter, 2002 All Rights Reserved iv TABLE OF CONTENTS Chapter 1. -

Type Designation for Cyclachaena Xanthiifolia (Euphrosyne Xanthiifolia) (Heliantheae, Asteraceae)
Phytotaxa 197 (2): 132–138 ISSN 1179-3155 (print edition) www.mapress.com/phytotaxa/ PHYTOTAXA Copyright © 2015 Magnolia Press Article ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.197.2.6 Type designation for Cyclachaena xanthiifolia (Euphrosyne xanthiifolia) (Heliantheae, Asteraceae) MANUEL B. CRESPO1*, CAROLINA PENA-MARTÍN2, ANNA SARAH BECKER3 & STEFAN DRESSLER3 1CIBIO (Instituto de la Biodiversidad) & dCARN, Universidad de Alicante, Apartado 99, E-03080 Alicante, Spain; e-mail: [email protected] 2Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Av. Alemania 01090, 4810101 Temuco, Chile; e-mail: [email protected] 3Senckenberg Forschungsinstitut, Herbarium Senckenbergianum Frankfurt/M. (FR), Senckenberganlage 25, D-60325 Frankfurt/M., Germany; e-mail: [email protected], [email protected] * Author for correspondence. Abstract Cyclachaena xanthiifolia (Iva xanthiifolia) is a North American member of the Heliantheae-Ambrosiinae (Asteraceae), which is currently accepted in the genus Euphrosyne. It is a nitrophilous plant, usually growing in disturbed grounds related to human activities, and this fact has probably favoured its introduction and naturalization in Eurasia and New Zealand. As part of the taxonomical work on the Iberian Compositae, typification of the name Cyclachaena xanthiifolia is effected here by selection of a lectotype and an epitype from the material housed at FR (Frankfurt am Main, Germany). The protologue and original material are discussed, and pictures of the relevant material are presented. Some historical aspects are also dis- cussed to justify typification. Key words: Ambrosiinae, Compositae, Cyclachaena, epitype, Euphrosyne, Iva, lectotype, nomenclature, typification Introduction The tribe Heliantheae (sensu Panero 2007) includes about 113 genera and ca.