The Birth of Classical Genetics As the Junction of Two Disciplines: Conceptual Change As Representational Change Marion Vorms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Models of the Gene Must Inform Data-Mining Strategies in Genomics
entropy Review Models of the Gene Must Inform Data-Mining Strategies in Genomics Łukasz Huminiecki Department of Molecular Biology, Institute of Genetics and Animal Biotechnology, Polish Academy of Sciences, 00-901 Warsaw, Poland; [email protected] Received: 13 July 2020; Accepted: 22 August 2020; Published: 27 August 2020 Abstract: The gene is a fundamental concept of genetics, which emerged with the Mendelian paradigm of heredity at the beginning of the 20th century. However, the concept has since diversified. Somewhat different narratives and models of the gene developed in several sub-disciplines of genetics, that is in classical genetics, population genetics, molecular genetics, genomics, and, recently, also, in systems genetics. Here, I ask how the diversity of the concept impacts data-integration and data-mining strategies for bioinformatics, genomics, statistical genetics, and data science. I also consider theoretical background of the concept of the gene in the ideas of empiricism and experimentalism, as well as reductionist and anti-reductionist narratives on the concept. Finally, a few strategies of analysis from published examples of data-mining projects are discussed. Moreover, the examples are re-interpreted in the light of the theoretical material. I argue that the choice of an optimal level of abstraction for the gene is vital for a successful genome analysis. Keywords: gene concept; scientific method; experimentalism; reductionism; anti-reductionism; data-mining 1. Introduction The gene is one of the most fundamental concepts in genetics (It is as important to biology, as the atom is to physics or the molecule to chemistry.). The concept was born with the Mendelian paradigm of heredity, and fundamentally influenced genetics over 150 years [1]. -

Epigenetics Is Normal Science, but Don't Call It Lamarckian
Extended Abstract Volume 6:3, 2020 DOI: 6.2472/imedipce.2020.6.002 Journal of Clinical Epigenetics ISSN: 2472-1158 Open Access Epigenetics is Normal Science, But Don’t Call It Lamarckian David Penny New Zealand Extended Abstract they are general (but not necessarily universal) because they are found in Excavates, which many people consider are ancestral to other eukaryotes [6, 7]. Similarly, an important point is that Epigenetic mechanisms are increasingly being accepted as an integral methylation, phosphorylation, etc. of the DNA and of the histones is a part of normal science. Earlier genetic studies appear to have part of normal evolution (at least in eukaryotes that have the assumed that the only changes were ‘mutations’ as a result of a nucleosome structure) [8]. One of the most interesting aspects is change in the DNA sequence. If any such changes altered the amino methylation (and hydroxymethylation) of cytosine, and this is found in acid sequences then it was effectively a mutation, and if it occurred in many deep branching eukaryotes [9]. Typically there is methylation of a germline cell (including in unicellular organisms) then it could be cytosine molecules, especially in CpG (cytosine followed by guanine) passed on to subsequent generations. However, it is increasingly regions. The role of small RNAs in helping regulate the levels of apparent that other changes are also important, for example, changes protein expression is discussed in [10]. The main point here is that to methylation patterns, and to the histone proteins. Epigenetics was protein expression levels are very variable in all cells – it is not just sometimes consider ‘additional’ to classical genetics, but it is still fully classical genetics that is important; there are many epigenetic dependent on DNA, RNA and protein sequences, and at least since [1] mechanisms affecting the differences in protein expression. -

Using Classical Genetics Simulator (CGS) to Teach Students the Basics of Genetic Research
Tested Studies for Laboratory Teaching Proceedings of the Association for Biology Laboratory Education Vol. 33, 85–94, 2012 Using Classical Genetics Simulator (CGS) to Teach Students the Basics of Genetic Research Jean Heitz1, Mark Wolansky2, and Ben Adamczyk3 1University of Wisconsin, Zoology Department, 250 N. Mills Street, Madison WI 53706 USA 2University of Alberta, Department of Biological Science, Edmonton AB, CAN T6G 2E9 3Genedata, MA, A-1 Cranberry HI, Lexington MA 02421 USA ([email protected];[email protected];[email protected]) Some experiments are not well suited to large introductory labs because of space, time and funding constraints. Among these are classical Mendelian genetics investigations. Cyber labs can overcome these constraints. Classi- cal Genetics Simulator (CGS) gives students the opportunity to perform controlled crosses with model organisms much like a geneticist would do. CGS provides populations of Drosophila, Arabidopsis or mice with unknown patterns of inheritance and gives students the tools to design and perform experiments to discover these patterns. Students require an understanding of the what, why and how of solving genetic problems to successfully complete our cyber genetics lab activities. Keywords: Mendelian genetics, simulator, classical genetics simulator (CGS), linkage, monohybrid, dihybrid Introduction In large introductory biology classes some labs are dif- How can students get practice doing what geneticists ficult to do because of space, time and funding constraints. actually do? Among these are classical Mendelian genetics investiga- An obvious way for students to get practice doing what tions. As a result, we have turned to cyber labs. Working geneticists do is to conduct actual crosses. -

The Sustained Impact of Model Organisms—In Genetics and Epigenetics
| COMMENTARY The Sustained Impact of Model Organisms—in Genetics and Epigenetics Nancy M. Bonini*,†,1 and Shelley L. Berger‡,1 *Department of Cellular and Developmental Biology, Perlman School of Medicine, †Department of Biology, and ‡Department of Genetics, University of Pennsylvania, Philadelphia, Pennsylvania 19104 KEYWORDS classical genetics; epigenetics; behavior; genomics; human disease Now that DNA sequencing can reveal disease-relevant muta- The Rise of Model Organism Genetics and the tions in humans, it is appropriate to consider the history of Embrace of Molecular Genetics genetic research on model organisms, and whether model Model organism genetics has revealed many fundamental organisms will continue to play an important role. Our view is biological processes. This was achieved by applying unbiased that genetic research in model organisms has had an in- genetic screens to reveal genes and their mechanisms of disputably enormous impact on our understanding of com- action in processes such as cell cycle control (Nasmyth plex biological pathways and their epigenetic regulation, and 2001), basic body plan specification and mechanisms of de- on the development of tools and approaches that have been velopment (Lewis 1978; Nusslein-Volhard and Wieschaus crucial for studies of human biology. We submit that model 1980), and cell death pathways (Ellis and Horvitz 1986; organisms will remain indispensable for interpreting complex Avery and Horvitz 1987), all of which were recognized with genomic data, for testing hypotheses regarding function, and Nobel prizes. for further study of complex behaviors, to reveal not only The sequencing of genomes revealed the breadth of the novel genetic players, but also the profound impact of epige- conservation of genes, and of entire pathways. -

Laboratory III Linkage
Genetics Laboratory III Biology 303 Spring 2007 Linkage Dr. Wadsworth Introduction same chromosome, the meiotic assortment of chromosomes alone would not predict the Genes are arranged as units in a linear order independent assortment of genes on those along chromosomes. The position of each gene chromosomes. In the absence of gene along the chromosome is called its locus. Each assortment, we would predict that the progeny gene has a unique chromosomal position and could only have the allelic combinations that therefore a unique locus. In fact, geneticists were present in the parent chromosomes. Genes often use the term genetic locus interchangeably that do not assort are said to show complete with the term gene. linkage. Alternatively, genes that independently assort are said to be unlinked. For the following The relative position of genes along a two examples, compare the genotypes and chromsomes can be analyzed by determining phenotypes generated in a dihybrid crosses of their linkage relationship. The concept of unlinked and completely linked genes. genetic linkage is outlined in this handout and will be discussed in class in the coming weeks. The position of a gene on a chromosome can be Example 1: A dihybrid cross for genes A and B, establish its identity. two unlinked genes. Generation Genotypes Phenotypes Specific Goals P1 AABBXaabb AB and ab 1. Conduct appropriate genetic crosses with a F1 AaBb AB mutant D. melanogaster to determine the following: F2 1AABB:2AABb: 9AB:3Ab:3aB:1ab 2AaBB:4AaBb:1AAbb: 2Aabb:1aaBB:2aaBb:1aabb a. Segregation b. Independent Assortment/Linkage Phenotypic ratio from dihybrid cross for c. -

G14TBS Part II: Population Genetics
G14TBS Part II: Population Genetics Dr Richard Wilkinson Room C26, Mathematical Sciences Building Please email corrections and suggestions to [email protected] Spring 2015 2 Preliminaries These notes form part of the lecture notes for the mod- ule G14TBS. This section is on Population Genetics, and contains 14 lectures worth of material. During this section will be doing some problem-based learning, where the em- phasis is on you to work with your classmates to generate your own notes. I will be on hand at all times to answer questions and guide the discussions. Reading List: There are several good introductory books on population genetics, although I found their level of math- ematical sophistication either too low or too high for the purposes of this module. The following are the sources I used to put together these lecture notes: • Gillespie, J. H., Population genetics: a concise guide. John Hopkins University Press, Baltimore and Lon- don, 2004. • Hartl, D. L., A primer of population genetics, 2nd edition. Sinauer Associates, Inc., Publishers, 1988. • Ewens, W. J., Mathematical population genetics: (I) Theoretical Introduction. Springer, 2000. • Holsinger, K. E., Lecture notes in population genet- ics. University of Connecticut, 2001-2010. Available online at http://darwin.eeb.uconn.edu/eeb348/lecturenotes/notes.html. • Tavar´eS., Ancestral inference in population genetics. In: Lectures on Probability Theory and Statistics. Ecole d'Et´esde Probabilit´ede Saint-Flour XXXI { 2001. (Ed. Picard J.) Lecture Notes in Mathematics, Springer Verlag, 1837, 1-188, 2004. Available online at http://www.cmb.usc.edu/people/stavare/STpapers-pdf/T04.pdf Gillespie, Hartl and Holsinger are written for non-mathematicians and are easiest to follow. -

Mendelian Gene ” and the ” Molecular Gene ” : Two Relevant Concepts of Genetic Units V Orgogozo, Alexandre E
The ” Mendelian Gene ” and the ” Molecular Gene ” : Two Relevant Concepts of Genetic Units V Orgogozo, Alexandre E. Peluffo, Baptiste Morizot To cite this version: V Orgogozo, Alexandre E. Peluffo, Baptiste Morizot. The ” Mendelian Gene ” and the ”Molec- ular Gene ” : Two Relevant Concepts of Genetic Units. Virginie Orgogozo. Genes and Evolu- tion, 119, Elsevier, pp.1-26, 2016, Current Topics in Developmental Biology, 978-0-12-417194-7. 10.1016/bs.ctdb.2016.03.002. hal-01354346 HAL Id: hal-01354346 https://hal.archives-ouvertes.fr/hal-01354346 Submitted on 19 Aug 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. ARTICLE IN PRESS The “Mendelian Gene” and the “Molecular Gene”: Two Relevant Concepts of Genetic Units V. Orgogozo*,1, A.E. Peluffo*, B. Morizot† *Institut Jacques Monod, UMR 7592, CNRS-Universite´ Paris Diderot, Sorbonne Paris Cite´, Paris, France †Universite´ Aix-Marseille, CNRS UMR 7304, Aix-en-Provence, France 1Corresponding author: e-mail address: [email protected] Contents 1. Introduction 2 2. The “Mendelian Gene” and the “Molecular Gene” 5 3. Current Literature Often Confuses the “Mendelian Gene” and the “Molecular Gene” Concepts 7 4. -

Molecular Population Genetics
| FLYBOOK ECOLOGY AND EVOLUTION Molecular Population Genetics Sònia Casillas*,† and Antonio Barbadilla*,†,1 *Institut de Biotecnologia i de Biomedicina, and yDepartament de Genètica i de Microbiologia, Campus Universitat Autònoma de Barcelona (UAB), 08193 Cerdanyola del Vallès, Barcelona, Spain ORCID IDs: 0000-0001-8191-0062 (S.C.); 0000-0002-0374-1475 (A.B.) ABSTRACT Molecular population genetics aims to explain genetic variation and molecular evolution from population genetics principles. The field was born 50 years ago with the first measures of genetic variation in allozyme loci, continued with the nucleotide sequencing era, and is currently in the era of population genomics. During this period, molecular population genetics has been revolutionized by progress in data acquisition and theoretical developments. The conceptual elegance of the neutral theory of molecular evolution or the footprint carved by natural selection on the patterns of genetic variation are two examples of the vast number of inspiring findings of population genetics research. Since the inception of the field, Drosophila has been the prominent model species: molecular variation in populations was first described in Drosophila and most of the population genetics hypotheses were tested in Drosophila species. In this review, we describe the main concepts, methods, and landmarks of molecular population genetics, using the Drosophila model as a reference. We describe the different genetic data sets made available by advances in molecular technologies, and the theoretical developments fostered by these data. Finally, we review the results and new insights provided by the population genomics approach, and conclude by enumerating challenges and new lines of inquiry posed by in- creasingly large population scale sequence data. -
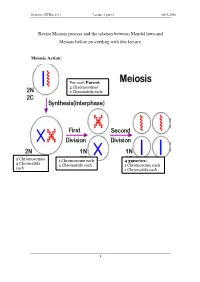
Revise Meiosis Process and the Relation Between Mendel Laws and Meiosis Before Proceeding with This Lecture
Genetics (BTBio 211) Lecture 3 part 2 2015-2016 Revise Meiosis process and the relation between Mendel laws and Meiosis before proceeding with this lecture. Meiosis Action: For each Parent 2 Chromosomes 2 Chromatids each 2 Chromosomes 1 Chromosome each 4 gametes: 4 Chromatid s 4 Chromatids each 1 Chromosome each each 1 Chromatids each 1 Genetics (BTBio 211) Lecture 3 part 2 2015-2016 LINKAGE AND CHROMOSOME MAPPING IN EUKARYOTES I. LINKAGE Genetic linkage is the tendency of genes that are located proximal to each other on a chromosome to be inherited together during meiosis. Genes whose loci are nearer to each other are less likely to be separated onto different chromatids during chromosomal crossover, and are therefore said to be genetically linked. Linked genes: Genes that are inherited together with other gene(s) in form of single unit as they are located on the same chromosome. For example: in fruit flies the genes for eye color and the genes for wing length are on the same chromosome, thus are inherited together. A couple of genes on chromosomes may be present either on the different or on the same chromosome. 1. The independent assortment of two genes located on different chromosomes. Mendel’s Law of Independent Assortment: during gamete formation, segregation of one gene pair is independent of other gene pairs because the traits he studied were determined by genes on different chromosomes. Consider two genes A and B, each with two alleles A a and B b on separate (different) chromosomes. 2 Genetics (BTBio 211) Lecture 3 part 2 2015-2016 Gametes of non-homologous chromosomes assort independently at anaphase producing 4 different genotypes AB, ab, Ab and aB with a genotypic ratio 1:1:1:1. -

The Hardy-Weinberg Equilibrium: Some Helpful Suggestions
the Hardy-WeinbergEquilibrium-Some0 Helpful Suggestions MaryT. Ortiz LorettaTaras Anthea M. Stavroulakis TN college General Biology curricula, the topic of Hardy-Weinberg equilibrium. Our experiences in classical genetics is typically followed by consider- teaching Mendelian probabilities and the Hardy- ation of population and evolutionary genetics. Weinberg model confirm that students enrolled in After fundamental genetic principles are studied, GeneralBiology who have not attainedthe established students are introduced to the Hardy-Weinbergequi- level of proficiency in mathematics have difficulty librium as a continuum of Mendelian Genetics in a participating in class discussions on this topic and population context. Population genetics provides a solving homework problems and examination Downloaded from http://online.ucpress.edu/abt/article-pdf/62/1/20/49339/4450821.pdf by guest on 28 September 2021 logical segue to the study of evolution. Populations questions. adhering to the Hardy-Weinbergequilibrium provide Mendel's laws provide the foundation for discus- a reference for studying evolutionary changes sions of evolutionary and population genetics (Dob- (Journet1986). With this as a startingpoint, Darwin's zhansky et al. 1997).The Hardy-Weinbergequilibrium observations can be justified and quantified, as well is a continuationof Mendel's principles, which show as expanded upon, to include additional aspects of that gene frequenciesunder certainconditions remain population and evolutionary genetics. constant from one generation to the next, thereby The Hardy-Weinbergequilibrium is an algebraic maintaining the allelic composition of the gene pool mathematical tool for predicting allele frequencies, of the population. During our presentation of the phenotypes and genotypes in populations (Cum- Hardy-Weinbergequilibrium, we link Punnettsquares mings 1997; Lewis 1997; Mader 1996;Postlethwait & and probabilitycalculations to previous discussions of Hopson 1995).Mertens (1992)proposed strategies for classicalinheritance. -

A Century of Geneticists Mutation to Medicine a Century of Geneticists Mutation to Medicine
A Century of Geneticists Mutation to Medicine http://taylorandfrancis.com A Century of Geneticists Mutation to Medicine Krishna Dronamraju CRC Press Taylor & Francis Group 6000 Broken Sound Parkway NW, Suite 300 Boca Raton, FL 33487-2742 © 2019 by Taylor & Francis Group, LLC CRC Press is an imprint of Taylor & Francis Group, an Informa business No claim to original U.S. Government works Printed on acid-free paper International Standard Book Number-13: 978-1-4987-4866-7 (Paperback) International Standard Book Number-13: 978-1-138-35313-8 (Hardback) This book contains information obtained from authentic and highly regarded sources. Reasonable efforts have been made to publish reliable data and information, but the author and publisher cannot assume responsibility for the validity of all materials or the consequences of their use. The authors and publishers have attempted to trace the copyright holders of all material reproduced in this publication and apologize to copyright holders if permission to publish in this form has not been obtained. If any copyright material has not been acknowledged please write and let us know so we may rectify in any future reprint. Except as permitted under U.S. Copyright Law, no part of this book may be reprinted, reproduced, trans- mitted, or utilized in any form by any electronic, mechanical, or other means, now known or hereafter invented, including photocopying, microfilming, and recording, or in any information storage or retrieval system, without written permission from the publishers. For permission to photocopy or use material electronically from this work, please access www.copyright .com (http://www.copyright.com/) or contact the Copyright Clearance Center, Inc. -

00326673.Pdf
Unda_standing Inheritance ~ hat like begets like–that what is now called a species begets offspring ;! of the same species—must have been evident to the earliest humans. ;! Recognition of the inheritance of variations within a species must also j; have come early, since domestication of animals undoubtedly involved II !-i elimination of individuals with undesirable characteristics (a penchant for human flesh, for example). The first animals to be domesticated may well have been members of the dog family, which were used as food, and domestication of canines may have started even before the advent of Homo” sapiens. The remains of an old hominid relative of ours, Hmno erectus (also known as Java or Peking man), have been found associated with those of a dog-like animal in 500,000-year-old fossils. The earliest canine remains associated with our own species are a mere 12,000 years old. The domestication of food plants probably began between 8000 and 9000 years ago, although some authorities contend that the domestication of cereals preceded that of most animals. Humans must also have very early related treating between “male” and “female” animals, including humans, with the subsequent issuance of offspring. Sexual repro- duction in plants was probably recognized much later—many plants, after all, are discreet] y bisexual—but at least 4000 years ago, as evidenced by the Babylonians’ selective breeding, through controlled pollination, of the date palm (Ph~jeni.~ du(fylif- eru),which occurs as separate male and female trees. (The dates borne by a female tree result from fertilization of its eggs by spern-containing pollen from male trees.) The oldest recorded thoughts about heredity appear in the religious writings of the ancient Hindus and Jews, which reveal recognition of the heritability of disease, health, and mental and physical characteristics.