Atypical Estuaries in NSW: Implications for Management of Lake Wollumboola
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

LOCALITY MAP Compartment 720 Nullica State Forest No.545 SOUTHERN REGION: EDEN MANAGEMENT AREA BOGGY CREEK Scale: 1:100,000
Bournda NR LOCALITY MAP Compartment 720 Nullica State Forest No.545 SOUTHERN REGION: EDEN MANAGEMENT AREA BOGGY CREEK Scale: 1:100,000 MERIMBULA LAKE Á Pambula ! Ben Boyd NP! Á Á Dobbyns Road PAMBULA RIVER P" YOWAKA RIVER G PAMBULA LAKE 720 Egan Peaks NR South East Forest NP PALESTINE CREEK CURALO LAGOON Eden ! Towns & Localities ! Sealed Road Major Rivers® Major Forest Road COCORA LAGOON State Forest National Parks SHADRACHS CREEK Planning Unit Formal Reserve Vacant CrownLand Informal Reserve NonForest Waterbodies Freehold NULLICA RIVER G Emergency Meeting Point Á Evacuation Route LEOS CREEK REEDY CREEK Haulage Route P" Helicopter Landing Site Á BOYDTOWN CREEK TOWAMBA RIVER Mount Imlay NP Prepared By: AndrewKemsley Harvest Plan Operational Map Compartment: 720 Version: 1 .................RE....G.I...O.NA.....L... M....ANA.........G.E...R.... A.PP.....R...O....V.AL................... State Forest: Nullica No: 545 APPROVED: DANIEL TUAN SOUTHERN REGION - Native Forests ³ DATE: 05/07/2012 Map Sheet: EDEN 8824-2S 45 46 47 A X 05 05 ^! ^ XX XX JA ^ CH # 720-3 Rd H B H 0# 3 HHS3 2 D 0# ú G B 0#0# H BB 1 720-6 Rd S2 BB 04 ú FH ^ 04 H L ^ J XX ^! KH ú E 4 0# S1 0# ^! JH B # úC1 B B É BB I J XX 03 745000E 46 47 BOUNDARIES NONHARVEST AREA FAUNA FEATURES ÉÉÉÉÉÉCompartment Boundary Special Management - FMZ 2 A PowerfulOwl ÉÉÉÉÉÉCoupe Boundary (100m either side) ^ Gang Gang Cockatoo Smoky Mouse Exclusion Area ^! Smoky Mouse ROADS Ridge & HeadwaterHabitat (80m) X Yellow-bellied Glider Major Forest # 32> Excluded Forest Varied Sittella Minor Forest Rocky Outcrop (0.1-0.5 ha, 20m) ^ Glossy Black-Cockatoo EPL Standard Existing (Major) X EPL Standard Existing (Minor) Cliff and buffer (20m) X Yellow-bellied Glider (Heard) EPL Licenced (New Construction) Slopes >30 (IHL4) ^ Eastern Pigmy Possum DRAINAGE FEATURE PROTECTION (EPL DUMPS & CROSSINGS FLORA FEATURES IHL 2 & TSL). -

Challenges of Saltmarsh Rehabilitation in a Non-Tidal Estuary – Perspectives on Delivery of On-Ground Works
CHALLENGES OF SALTMARSH REHABILITATION IN A NON-TIDAL ESTUARY – PERSPECTIVES ON DELIVERY OF ON-GROUND WORKS M Barnett, N Dixon, A Halcrow, S Tsaprounis, D Ryan Wyong Shire Council, Wyong NSW Background Saltmarshes are a unique component of coastal estuaries in Australia that exist at the land and water interface. Conditions in these areas are highly dynamic and biota must be adapted and specialised to cope with varying degrees of salinity, tidal inundation, competition and disturbance. Their unique position allows saltmarshes to function in several ways including flood and erosion control, storm surge buffering, water quality improvement and seagrass wrack assimilation. For these and many other reasons, not the least being biodiversity, saltmarshes need to be conserved as vital parts of the Australian coastline. Saltmarshes are a diminishing habitat in the coastal biosphere of Australia. New South Wales in particular has the least amount of coastal saltmarsh remaining of any state. Saltmarsh occurs along the entire coastline of NSW, but is generally limited to small patches that have been highly modified. In recognition of the decline and uniqueness of saltmarsh in NSW, it has been listed as an Endangered Ecological Community in NSW under the Threatened Species Conservation Act. On the local scale, it is believed that approximately 85% of the saltmarshes around Tuggerah Lakes have been destroyed, degraded or fragmented since European settlement. Saltmarsh recovery on rehabilitated sites can occur naturally, be assisted through active planting and/or bush regeneration or a combination of both. Natural recovery of saltmarsh communities occurs after disturbance via establishment of seedlings or vegetative expansion of remaining plants. -

NSW Vagrant Bird Review
an atlas of the birds of new south wales and the australian capital territory Vagrant Species Ian A.W. McAllan & David J. James The species listed here are those that have been found on very few occasions (usually less than 20 times) in NSW and the ACT, and are not known to have bred here. Species that have been recorded breeding in NSW are included in the Species Accounts sections of the three volumes, even if they have been recorded in the Atlas area less than 20 times. In determining the number of records of a species, when several birds are recorded in a short period together, or whether alive or dead, these are here referred to as a ‘set’ of records. The cut-off date for vagrant records and reports is 31 December 2019. As with the rest of the Atlas, the area covered in this account includes marine waters east from the NSW coast to 160°E. This is approximately 865 km east of the coast at its widest extent in the south of the State. The New South Wales-Queensland border lies at about 28°08’S at the coast, following the centre of Border Street through Coolangatta and Tweed Heads to Point Danger (Anon. 2001a). This means that the Britannia Seamounts, where many rare seabirds have been recorded on extended pelagic trips from Southport, Queensland, are east of the NSW coast and therefore in NSW and the Atlas area. Conversely, the lookout at Point Danger is to the north of the actual Point and in Queensland but looks over both NSW and Queensland marine waters. -
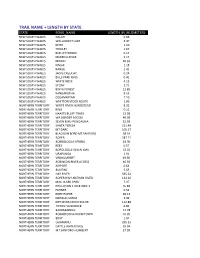
Trail Name + Length by State
TRAIL NAME + LENGTH BY STATE STATE ROAD_NAME LENGTH_IN_KILOMETERS NEW SOUTH WALES GALAH 0.66 NEW SOUTH WALES WALLAGOOT LAKE 3.47 NEW SOUTH WALES KEITH 1.20 NEW SOUTH WALES TROLLEY 1.67 NEW SOUTH WALES RED LETTERBOX 0.17 NEW SOUTH WALES MERRICA RIVER 2.15 NEW SOUTH WALES MIDDLE 40.63 NEW SOUTH WALES NAGHI 1.18 NEW SOUTH WALES RANGE 2.42 NEW SOUTH WALES JACKS CREEK AC 0.24 NEW SOUTH WALES BILLS PARK RING 0.41 NEW SOUTH WALES WHITE ROCK 4.13 NEW SOUTH WALES STONY 2.71 NEW SOUTH WALES BINYA FOREST 12.85 NEW SOUTH WALES KANGARUTHA 8.55 NEW SOUTH WALES OOLAMBEYAN 7.10 NEW SOUTH WALES WHITTON STOCK ROUTE 1.86 NORTHERN TERRITORY WAITE RIVER HOMESTEAD 8.32 NORTHERN TERRITORY KING 0.53 NORTHERN TERRITORY HAASTS BLUFF TRACK 13.98 NORTHERN TERRITORY WA BORDER ACCESS 40.39 NORTHERN TERRITORY SEVEN EMU‐PUNGALINA 52.59 NORTHERN TERRITORY SANTA TERESA 251.49 NORTHERN TERRITORY MT DARE 105.37 NORTHERN TERRITORY BLACKGIN BORE‐MT SANFORD 38.54 NORTHERN TERRITORY ROPER 287.71 NORTHERN TERRITORY BORROLOOLA‐SPRING 63.90 NORTHERN TERRITORY REES 0.57 NORTHERN TERRITORY BOROLOOLA‐SEVEN EMU 32.02 NORTHERN TERRITORY URAPUNGA 1.91 NORTHERN TERRITORY VRDHUMBERT 49.95 NORTHERN TERRITORY ROBINSON RIVER ACCESS 46.92 NORTHERN TERRITORY AIRPORT 0.64 NORTHERN TERRITORY BUNTINE 5.63 NORTHERN TERRITORY HAY RIVER 335.62 NORTHERN TERRITORY ROPER HWY‐NATHAN RIVER 134.20 NORTHERN TERRITORY MAC CLARK PARK 7.97 NORTHERN TERRITORY PHILLIPSON STOCK ROUTE 55.84 NORTHERN TERRITORY FURNER 0.54 NORTHERN TERRITORY PORT ROPER 40.13 NORTHERN TERRITORY NDHALA GORGE 3.49 NORTHERN TERRITORY -

Walk-Issue14-1963.Pdf
1963 Terms and Conditions of Use Copies of Walk magazine are made available under Creative Commons - Attribution Non-Commercial Share Alike copyright. Use of the magazine. You are free: • To Share- to copy, distribute and transmit the work • To Remix- to adapt the work Under the following conditions (unless you receive prior written authorisation from Melbourne Bushwalkers Inc.): • Attribution- You must attribute the work (but not in any way that suggests that Melbourne Bushwalkers Inc. endorses you or your use of the work). • Noncommercial- You may not use this work for commercial purposes. • Share Alike- If you alter, transform, or build upon this work, you may distribute the resulting work only under the same or similar license to this one. Disclaimer of Warranties and Limitations on Liability. Melbourne Bushwalkers Inc. makes no warranty as to the accuracy or completeness of any content of this work. Melbourne Bushwalkers Inc. disclaims any warranty for the content, and will not be liable for any damage or loss resulting from the use of any content. ----···············------------------------------· • BUSHWALKING • CAVING • ROCK CLIMBING • CAMPING • SKI TOURING PROVIDE A CHALLENGE TO MAN AND HIS EQUIPMENT, FOR OVER 30 YEARS, PADDYMADE CAMP GEAR HAS PROVED ITS WORTH TO THOUSANDS OF WALKERS AND OUT-OF-DOORS ADVEN TURERS. MAKE SURE YOU, TOO, HAVE THE BEST OF GEAR. From- PADDY PALLIN Py. ltd. 201 CASTLEREAGH STREET, SYDNEY - Phone BM 2685 Ask for our Latest Price List Get your copy of "Bushwalking - --- and Camping," by Paddy Pallin -5/6 posted --------------------------------------------------· CWalk A JOURNAL OF THE MELBOURNE BUSHW ALKERS NUMBER FOURTEEN 1963 CONTENTS: * BY THE PEOPLE 'l ... -

Assessing Estuary Ecosystem Health: Sampling, Data Analysis and Reporting Protocols
Assessing estuary ecosystem health: Sampling, data analysis and reporting protocols NSW Natural Resources Monitoring, Evaluation and Reporting Program Cover image: Meroo Lake, Meroo National Park/M Jarman OEH © 2013 State of NSW and Office of Environment and Heritage With the exception of photographs, the State of NSW and Office of Environment and Heritage are pleased to allow this material to be reproduced in whole or in part for educational and non-commercial use, provided the meaning is unchanged and its source, publisher and authorship are acknowledged. Specific permission is required for the reproduction of photographs. The Office of Environment and Heritage (OEH) has compiled this publication in good faith, exercising all due care and attention. No representation is made about the accuracy, completeness or suitability of the information in this publication for any particular purpose. OEH shall not be liable for any damage which may occur to any person or organisation taking action or not on the basis of this publication. Readers should seek appropriate advice when applying the information to their specific needs. Published by: Office of Environment and Heritage 59 Goulburn Street, Sydney NSW 2000 PO Box A290, Sydney South NSW Phone: (02) 9995 5000 (switchboard) Phone: 131 555 (environment information and publications requests) Phone: 1300 361 967 (national parks, general environmental inquiries and publications requests) Fax: (02) 9995 5999 TTY users: phone 133 677, then ask for 131 555 Speak and listen users: phone 1300 555 727, -

Entrance Dynamics and Beach Condition at the Entrance and North Entrance Beaches
Wyong Shire Council Entrance Dynamics and Beach Condition at The Entrance and North Entrance Beaches April 2011 Entrance Dynamics and Beach Condition at The Entrance and North Entrance Beaches Prepared by Umwelt (Australia) Pty Limited on behalf of Wyong Council Project Director: Pam Dean-Jones Project Manager: Pam Dean-Jones Report No. 1869/RO5/Final Date: APRIL 2011 2/20 The Boulevarde PO Box 838 Toronto NSW 2283 Ph: 02 4950 5322 Fax: 02 4950 5737 Email: [email protected] Website: www.umwelt.com.au Entrance Dynamics Executive Summary Executive Summary This study was commissioned to provide further analysis of the sediment transport processes affecting sediment budget and coastal morphology at North Entrance Beach and The Entrance to Tuggerah Lake. The aim of the study was to clarify sediment transport linkages between The Entrance channel and North Entrance Beach and to identify and evaluate potential options for managing sedimentary processes in this dynamic coastal landscape, to reduce risks associated with coastal erosion hazards. North Entrance Beach lies at the southern end of the zeta curved Tuggerah coastal embayment. The southern extension of the beach is an intermittent spit and berm which forms at the mouth of the Tuggerah lake entrance channel. In historical times the mouth of the estuary closed for periods of up to three years. The sand deposits were scoured out of the entrance area by major flood flows. Since dredging of the outer entrance shoals commenced in 1993, the average channel width has declined compared to the average pre dredging condition. The range of channel conditions has also narrowed. -

Environmental Sensitivity of Lake Wollumboola P a G E | 1
Environmental Sensitivity of Lake Wollumboola P a g e | 1 Environmental Sensitivity of Lake Wollumboola: Input to Considerations of Development Applications for Long Bow Point, Culburra Prepared by: Dr Peter Scanes, Dr Angus Ferguson, Jaimie Potts Estuaries and Catchments Science NSW Office of Environment and Heritage Environmental Sensitivity of Lake Wollumboola P a g e | 2 Environmental Sensitivity of Lake Wollumboola Input to Considerations of Development Applications for Long Bow Point, Culburra Executive Summary ............................................................................................................... 3 OEH analysis of the ecological and biogeochemical functioning of Lake Wollumboola ....... 3 OEH assessment of claims made by Realty Realizations ..................................................... 5 General implications of this analysis for development around Lake Wollumboola ............. 6 Introduction .......................................................................................................................... 7 Is Lake Wollumboola Like All other Estuaries? ....................................................................... 8 How does Lake Wollumboola Function? .............................................................................. 13 Conceptual model of lake function .................................................................................. 13 Entrance opening and salinity regime .............................................................................. 15 Stratification -

Ecological Studies on Illawarha Lake
ECOLOGICAL STUDIES ON ILLAWARHA LAKE WITH SPECIAL REFERENCE TO Zostera capricorni Ascherson. By Malcolm McD. Harris, B.A. ( Univ. of New England, Armidale ) A thesis submitted in fulfilment of the requirements for the degree of Master of Science of the University of New South Wales. School of Botany, University of New South Wales. January, 1977 UNIVERSITY OF N.S.W. 19851 16 SEP. 77 LIBRARY THIS IS TO CERTIFY that the work described in this thesis has not been submitted for a higher degree at any other university or institution. (iii) SUMMARY This thesis describes aspects of the ecology of Illawarra Lake, with special reference to the biology of the seagrass, Zostera capricomi Aschers. Observations were made from the air, from power boats, by wading and by SCUBA diving, over the period 1972 - 1976. Use has also been made of aerial photographs. The environmental factors studied include both sediment characteristics and water quality. Correlation coefficients have been calculated and used in the assessment of the functional relationships between the parameters examined. Reference has been made to corroborative evidence from a number of sources. The relationship between the distribution and biomass of the benthic flora of Illawarra Lake, and the selected environmental parameters, is examined. Seven other coastal saline lagoons were observed so that observations made and the conclusions drawn for Illawarra Lake, could be seen in the wider context. Long term observations and analyses have been made of the morphology, growth and flowering cycles of Z. capricomi. Evidence is presented showing some diagnostic features,used in published accounts to distinguish between Z. -

NPWS Annual Report 2000-2001 (PDF
Annual report 2000-2001 NPWS mission NSW national Parks & Wildlife service 2 Contents Director-General’s foreword 6 3 Conservation management 43 Working with Aboriginal communities 44 Overview 8 Joint management of national parks 44 Mission statement 8 Performance and future directions 45 Role and functions 8 Outside the reserve system 46 Partners and stakeholders 8 Voluntary conservation agreements 46 Legal basis 8 Biodiversity conservation programs 46 Organisational structure 8 Wildlife management 47 Lands managed for conservation 8 Performance and future directions 48 Organisational chart 10 Ecologically sustainable management Key result areas 12 of NPWS operations 48 Threatened species conservation 48 1 Conservation assessment 13 Southern Regional Forest Agreement 49 NSW Biodiversity Strategy 14 Caring for the environment 49 Regional assessments 14 Waste management 49 Wilderness assessment 16 Performance and future directions 50 Assessment of vacant Crown land in north-east New South Wales 19 Managing our built assets 51 Vegetation surveys and mapping 19 Buildings 51 Wetland and river system survey and research 21 Roads and other access 51 Native fauna surveys and research 22 Other park infrastructure 52 Threat management research 26 Thredbo Coronial Inquiry 53 Cultural heritage research 28 Performance and future directions 54 Conservation research and assessment tools 29 Managing site use in protected areas 54 Performance and future directions 30 Performance and future directions 54 Contributing to communities 55 2 Conservation planning -

South Coast Shorebird Recovery Newsletter 2017 18
SHOREBIRD RECOVERY NEWSLETTER south coast 2017/18 Season This Season in Shorebirds early raven predation dispersed the nesting birds across five sites. The volunteers really stepped up to the chal- The South Coast Shorebird Recovery Program contin- lenge and we achieved at least 20 fledglings for 2017/18. ues to monitor and protect endangered nesting shore- birds along our beaches, train and support volunteers The endangered Pied Oystercatcher breeding pair num- along the south coast and obtain funding to undertake bers were similar to last season, 27 pairs were monitored comprehensive fox control and community education with nests. A few more breeding pairs are quite remote, so programs. The program has been running now for al- there were likely a few additional nests that were missed. most 20 years and the ongoing support on the south But we will get better with time and more vols! Overall coast is inspiring! the Pied Oystercatchers fledged 17 chicks this season. This season the amazing 92 volunteers monitored a total Thanks to Saving our Species, Crown Lands and the Na- of 118 endangered shorebird nests between Gerroa and tional Parks and Wildlife Service our comprehensive fox Batemans Bay and recorded 3563 volunteer hours. Their control program was maintained in 2017/18 to reduce passion and dedication to shorebird recovery keeps the predation of shorebird eggs and chicks. The Local Land program running and has immeasurable effects through Services and Holiday Havens funded yet another awesome their links into the local community. These birds nest right season of community shorebird education programs. on our beaches so raising awareness, improving knowledge and local community support are extremely The ongoing support for our local shorebird recovery important for their survival into the future. -

The Entrance Morphodynamic Modelling Prepared for NSW Office of Environment and Heritage
Tuggerah Lakes– The Entrance Morphodynamic Modelling Prepared for NSW Office of Environment and Heritage Tuggerah Lakes – The Entrance Morphodynamic Modelling Prepared for NSW Office of Environment and Heritage Cardno (NSW/ACT) Pty Ltd ABN 95 001 145 035 Level 9 The Forum 203 Pacific Highway St Leonards NSW 2065 Australia Telephone: 02 9496 7700 Facsimile: 02 9439 5170 International: +61 2 9496 7700 [email protected] www.cardno.com.au © State of NSW and Office of Environment and Heritage November 2013 This document is produced by Cardno solely for the benefit and use by the client in accordance with the terms of the engagement. Cardno does not and shall not assume any responsibility or liability whatsoever to any third party arising out of any use or reliance by any third party on the content of this document. Tuggerah Lakes – The Entrance Morphodynamic Modelling Prepared for NSW Office of Environment and Heritage EXECUTIVE SUMMARY The Tuggerah Lakes system consists of three inter-connected shallow coastal lakes (Tuggerah, Budgewoi and Munmorah) that have a weak and intermittent connection to the ocean at The Entrance. The lakes have a history of water quality issues with development of their catchments leading to increased nutrient, and to a lesser extent sediment, loads. The lakes’ entrance is currently managed through regular dredging by Wyong Council to ensure that the entrance does not close and to achieve objectives related to flooding, water quality in the lower entrance channels and recreational amenity. The dredging does not affect the general water quality in the lakes. Previous assessments have considered the possibility of constructing a training wall or training walls at the entrance to Tuggerah Lakes with the objective of maintaining a permanent entrance to the lakes and thus improving water flows and water quality.