Habitability on Early Mars and the Search for Biosignatures with the Exomars Rover
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Global Structure of the Martian Dichotomy: an Elliptical Impact Basin? J
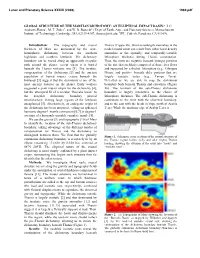
Lunar and Planetary Science XXXIX (2008) 1980.pdf GLOBAL STRUCTURE OF THE MARTIAN DICHOTOMY: AN ELLIPTICAL IMPACT BASIN? J. C. Andrews-Hanna1, M. T. Zuber1, and W. B. Banerdt2 (1Dept. of Earth, Atm., and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA 02139-4307, [email protected]; 2JPL, Caltech, Pasadena, CA 91109). Introduction: The topography and crustal Tharsis (Figure 2b). Short-wavelength anomalies in the thickness of Mars are dominated by the near- model crustal roots can result from either local density hemispheric dichotomy between the southern anomalies or the spatially- and temporally-variable highlands and northern lowlands. The dichotomy lithosphere thickness during Tharsis construction. boundary can be traced along an apparently irregular Thus, the roots are negative beneath younger portions path around the planet, except where it is buried of the rise that are likely composed of dense lava flows beneath the Tharsis volcanic rise [1]. The isostatic and supported by a thicker lithosphere (e.g., Olympus compensation of the dichotomy [2] and the ancient Mons), and positive beneath older portions that are population of buried impact craters beneath the largely isostatic today (e.g., Tempe Terra). lowlands [3] suggest that the dichotomy is one of the Nevertheless, we are able to map the dichotomy most ancient features on the planet. Early workers boundary both beneath Tharsis and elsewhere (Figure suggested a giant impact origin for the dichotomy [4], 2b). The location of the sub-Tharsis dichotomy but the attempted fit of a circular “Borealis basin” to boundary is largely insensitive to the choice of the irregular dichotomy boundary proved lithosphere thickness. -

Mars Reconnaissance Orbiter
Chapter 6 Mars Reconnaissance Orbiter Jim Taylor, Dennis K. Lee, and Shervin Shambayati 6.1 Mission Overview The Mars Reconnaissance Orbiter (MRO) [1, 2] has a suite of instruments making observations at Mars, and it provides data-relay services for Mars landers and rovers. MRO was launched on August 12, 2005. The orbiter successfully went into orbit around Mars on March 10, 2006 and began reducing its orbit altitude and circularizing the orbit in preparation for the science mission. The orbit changing was accomplished through a process called aerobraking, in preparation for the “science mission” starting in November 2006, followed by the “relay mission” starting in November 2008. MRO participated in the Mars Science Laboratory touchdown and surface mission that began in August 2012 (Chapter 7). MRO communications has operated in three different frequency bands: 1) Most telecom in both directions has been with the Deep Space Network (DSN) at X-band (~8 GHz), and this band will continue to provide operational commanding, telemetry transmission, and radiometric tracking. 2) During cruise, the functional characteristics of a separate Ka-band (~32 GHz) downlink system were verified in preparation for an operational demonstration during orbit operations. After a Ka-band hardware anomaly in cruise, the project has elected not to initiate the originally planned operational demonstration (with yet-to-be used redundant Ka-band hardware). 201 202 Chapter 6 3) A new-generation ultra-high frequency (UHF) (~400 MHz) system was verified with the Mars Exploration Rovers in preparation for the successful relay communications with the Phoenix lander in 2008 and the later Mars Science Laboratory relay operations. -

Lessons from Insight for Future Planetary Seismology
Open Archive Toulouse Archive Ouverte (OATAO ) OATAO is an open access repository that collects the wor of some Toulouse researchers and ma es it freely available over the web where possible. This is a publisher's version published in: https://oatao.univ-toulouse.fr/26931 Official URL : https://doi.org/10.1029/2019JE006353 To cite this version : Panning, M. P. and Pike, W. T. and Lognonné, Philippe,... [et al.]On͈Deck Seismology: Lessons from InSight for Future Planetary Seismology. (2020) Journal of Geophysical Research: Planets, 125 (4). ISSN 2169-9097 Any correspondence concerning this service should be sent to the repository administrator: [email protected] RESEARCH ARTICLE On-Deck Seismology: Lessons from InSight for Future 10.1029/2019JE006353 Planetary Seismology Special Section: 1 2 3 1 4 5 InSightatMars M. P. Panning , W. T. Pike , P. Lognonné , W. B. Banerdt , N. Murdoch , D. Banfield , C. Charalambous2 , S. Kedar1, R. D. Lorenz6 , A. G. Marusiak7 , J. B. McClean2,8 ,C. 1 9 2 10 Key Points: Nunn , S. C. Stähler ,A.E.Stott , and T. Warren • Based on InSight recordings, 1 2 atmospheric noise is amplified for Jet Propulsion Laboratory, California Institute of Technology, Pasadena, CA, USA, Department of Electrical and seismic sensors on deck, consistent Electronic Engineering, Imperial College London, London, UK, 3Planetology and Space Science Team, Université de with Viking observations Paris, Institut de Physique du Globe, CNRS, Paris, France, 4Department of Electronics, Optronics and Signal Processing, • -

Gnc 2021 Abstract Book
GNC 2021 ABSTRACT BOOK Contents GNC Posters ................................................................................................................................................... 7 Poster 01: A Software Defined Radio Galileo and GPS SW receiver for real-time on-board Navigation for space missions ................................................................................................................................................. 7 Poster 02: JUICE Navigation camera design .................................................................................................... 9 Poster 03: PRESENTATION AND PERFORMANCES OF MULTI-CONSTELLATION GNSS ORBITAL NAVIGATION LIBRARY BOLERO ........................................................................................................................................... 10 Poster 05: EROSS Project - GNC architecture design for autonomous robotic On-Orbit Servicing .............. 12 Poster 06: Performance assessment of a multispectral sensor for relative navigation ............................... 14 Poster 07: Validation of Astrix 1090A IMU for interplanetary and landing missions ................................... 16 Poster 08: High Performance Control System Architecture with an Output Regulation Theory-based Controller and Two-Stage Optimal Observer for the Fine Pointing of Large Scientific Satellites ................. 18 Poster 09: Development of High-Precision GPSR Applicable to GEO and GTO-to-GEO Transfer ................. 20 Poster 10: P4COM: ESA Pointing Error Engineering -

MARS DURING the PRE-NOACHIAN. J. C. Andrews-Hanna1 and W. B. Bottke2, 1Lunar and Planetary La- Boratory, University of Arizona
Fourth Conference on Early Mars 2017 (LPI Contrib. No. 2014) 3078.pdf MARS DURING THE PRE-NOACHIAN. J. C. Andrews-Hanna1 and W. B. Bottke2, 1Lunar and Planetary La- boratory, University of Arizona, Tucson, AZ 85721, [email protected], 2Southwest Research Institute and NASA’s SSERVI-ISET team, 1050 Walnut St., Suite 300, Boulder, CO 80302. Introduction: The surface geology of Mars appar- ing the pre-Noachian was ~10% of that during the ently dates back to the beginning of the Early Noachi- LHB. Consideration of the sawtooth-shaped exponen- an, at ~4.1 Ga, leaving ~400 Myr of Mars’ earliest tially declining impact fluxes both in the aftermath of evolution effectively unconstrained [1]. However, an planet formation and during the Late Heavy Bom- enduring record of the earlier pre-Noachian conditions bardment [5] suggests that the impact flux during persists in geophysical and mineralogical data. We use much of the pre-Noachian was even lower than indi- geophysical evidence, primarily in the form of the cated above. This bombardment history is consistent preservation of the crustal dichotomy boundary, to- with a late heavy bombardment (LHB) of the inner gether with mineralogical evidence in order to infer the Solar System [6] during which HUIA formed, which prevailing surface conditions during the pre-Noachian. followed the planet formation era impacts during The emerging picture is a pre-Noachian Mars that was which the dichotomy formed. less dynamic than Noachian Mars in terms of impacts, Pre-Noachian Tectonism and Volcanism: The geodynamics, and hydrology. crust within each of the southern highlands and north- Pre-Noachian Impacts: We define the pre- ern lowlands is remarkably uniform in thickness, aside Noachian as the time period bounded by two impacts – from regions in which it has been thickened by volcan- the dichotomy-forming impact and the Hellas-forming ism (e.g., Tharsis, Elysium) or thinned by impacts impact. -

EDL – Lessons Learned and Recommendations
."#!(*"# 0 1(%"##" !)"#!(*"#* 0 1"!#"("#"#(-$" ."!##("""*#!#$*#( "" !#!#0 1%"#"! /!##"*!###"#" #"#!$#!##!("""-"!"##&!%%!%&# $!!# %"##"*!%#'##(#!"##"#!$$# /25-!&""$!)# %"##!""*&""#!$#$! !$# $##"##%#(# ! "#"-! *#"!,021 ""# !"$!+031 !" )!%+041 #!( !"!# #$!"+051 # #$! !%#-" $##"!#""#$#$! %"##"#!#(- IPPW Enabled International Collaborations in EDL – Lessons Learned and Recommendations: Ethiraj Venkatapathy1, Chief Technologist, Entry Systems and Technology Division, NASA ARC, 2 Ali Gülhan , Department Head, Supersonic and Hypersonic Technologies Department, DLR, Cologne, and Michelle Munk3, Principal Technologist, EDL, Space Technology Mission Directorate, NASA. 1 NASA Ames Research Center, Moffett Field, CA [email protected]. 2 Deutsches Zentrum für Luft- und Raumfahrt e.V. (DLR), German Aerospace Center, [email protected] 3 NASA Langley Research Center, Hampron, VA. [email protected] Abstract of the Proposed Talk: One of the goals of IPPW has been to bring about international collaboration. Establishing collaboration, especially in the area of EDL, can present numerous frustrating challenges. IPPW presents opportunities to present advances in various technology areas. It allows for opportunity for general discussion. Evaluating collaboration potential requires open dialogue as to the needs of the parties and what critical capabilities each party possesses. Understanding opportunities for collaboration as well as the rules and regulations that govern collaboration are essential. The authors of this proposed talk have explored and established collaboration in multiple areas of interest to IPPW community. The authors will present examples that illustrate the motivations for the partnership, our common goals, and the unique capabilities of each party. The first example involves earth entry of a large asteroid and break-up. NASA Ames is leading an effort for the agency to assess and estimate the threat posed by large asteroids under the Asteroid Threat Assessment Project (ATAP). -

Exploration of Mars by the European Space Agency 1
Exploration of Mars by the European Space Agency Alejandro Cardesín ESA Science Operations Mars Express, ExoMars 2016 IAC Winter School, November 20161 Credit: MEX/HRSC History of Missions to Mars Mars Exploration nowadays… 2000‐2010 2011 2013/14 2016 2018 2020 future … Mars Express MAVEN (ESA) TGO Future ESA (ESA- Studies… RUSSIA) Odyssey MRO Mars Phobos- Sample Grunt Return? (RUSSIA) MOM Schiaparelli ExoMars 2020 Phoenix (ESA-RUSSIA) Opportunity MSL Curiosity Mars Insight 2020 Spirit The data/information contained herein has been reviewed and approved for release by JPL Export Administration on the basis that this document contains no export‐controlled information. Mars Express 2003-2016 … First European Mission to orbit another Planet! First mission of the “Rosetta family” Up and running since 2003 Credit: MEX/HRSC First European Mission to orbit another Planet First European attempt to land on another Planet Original mission concept Credit: MEX/HRSC December 2003: Mars Express Lander Release and Orbit Insertion Collission trajectory Bye bye Beagle 2! Last picture Lander after release, release taken by VMC camera Insertion 19/12/2003 8:33 trajectory Credit: MEX/HRSC Beagle 2 was found in January 2015 ! Only 6km away from landing site OK Open petals indicate soft landing OK Antenna remained covered Lessons learned: comms at all time! Credit: MEX/HRSC Mars Express: so many missions at once Mars Mission Phobos Mission Relay Mission Credit: MEX/HRSC Mars Express science investigations Martian Moons: Phobos & Deimos: Ionosphere, surface, -

Page 1 E X O M a R S E X O M a R S
NOTE ADDED BY JPL WEBMASTER: This document was prepared by the European Space Agency. The content has not been approved or adopted by, NASA, JPL, or the California Institute of Technology. This document is being made available for information purposes only, and any views and opinions expressed herein do not necessarily state or reflect those of NASA, JPL, or the California Institute of Technology. EE XX OO MM AA RR SS ExoMars Status th J. L. Vago and the ExoMars Project Team 20 MEPAG Meeting 3–4 March 2009, Arlington, VA (USA) ExoMars Original Objectives Technology Demonstration Objectives : Entry, Descent, and Landing (EDL) of a large payload on the surface of Mars; Surface mobility with a rover having several kilometres range; Access to the subsurface with a drill to acquire samples down to 2 metres; Automatic sample preparation and distribution for analysis with scientific instruments. Scientific Objectives (in order of priority): To search for signs of past and present life on Mars; To characterise the water/geochemical environment as a function of depth in the shallow subsurface; To study the surface environment and identify hazards to future human missions; To investigate the planet’s subsurface and deep interior to better understand its evolution and habitability. What is ExoMars Now? KEY REQUIREMENTS FOR EXOMARS : (but also for all future ESA Mars exploration missions) Clear synergy of technology and science goals: ExoMars has to land; ExoMars has to rove; ExoMars has to drill; ExoMars has to perform novel organics -

Amino Acids and the Asymmetry of Life; Springer, 2008.Pdf
Amino Acids and the Asymmetry of Life Advances in Astrobiology and Biogeophysics springer.com This series aims to report new developments in research and teaching in the inter- disciplinary fields of astrobiology and biogeophysics. This encompasses all aspects of research into the origins of life – from the creation of matter to the emergence of complex life forms – and the study of both structure and evolution of planetary ecosystems under a given set of astro- and geophysical parameters. The methods con- sidered can be of theoretical, computational, experimental and observational nature. Preference will be given to proposals where the manuscript puts particular emphasis on the overall readability in view of the broad spectrum of scientific backgrounds involved in astrobiology and biogeophysics. The type of material considered for publication includes: • Topical monographs • Lectures on a new field, or presenting a new angle on a classical field • Suitably edited research reports • Compilations of selected papers from meetings that are devoted to specific topics The timeliness of a manuscript is more important than its form which may be un- finished or tentative. Publication in this new series is thus intended as a service to the international scientific community in that the publisher, Springer-Verlag, offers global promotion and distribution of documents which otherwise have a restricted readership. Once published and copyrighted, they can be documented in the scientific literature. Series Editors: Dr. André Brack Dr. Christopher P. McKay Centre de Biophysique Moléculaire NASA Ames Research Center CNRS, Rue Charles Sadron Moffet Field 45071 Orléans, Cedex 2, France CA 94035,USA [email protected] Dr. -

The Pancam Instrument for the Exomars Rover
ASTROBIOLOGY ExoMars Rover Mission Volume 17, Numbers 6 and 7, 2017 Mary Ann Liebert, Inc. DOI: 10.1089/ast.2016.1548 The PanCam Instrument for the ExoMars Rover A.J. Coates,1,2 R. Jaumann,3 A.D. Griffiths,1,2 C.E. Leff,1,2 N. Schmitz,3 J.-L. Josset,4 G. Paar,5 M. Gunn,6 E. Hauber,3 C.R. Cousins,7 R.E. Cross,6 P. Grindrod,2,8 J.C. Bridges,9 M. Balme,10 S. Gupta,11 I.A. Crawford,2,8 P. Irwin,12 R. Stabbins,1,2 D. Tirsch,3 J.L. Vago,13 T. Theodorou,1,2 M. Caballo-Perucha,5 G.R. Osinski,14 and the PanCam Team Abstract The scientific objectives of the ExoMars rover are designed to answer several key questions in the search for life on Mars. In particular, the unique subsurface drill will address some of these, such as the possible existence and stability of subsurface organics. PanCam will establish the surface geological and morphological context for the mission, working in collaboration with other context instruments. Here, we describe the PanCam scientific objectives in geology, atmospheric science, and 3-D vision. We discuss the design of PanCam, which includes a stereo pair of Wide Angle Cameras (WACs), each of which has an 11-position filter wheel and a High Resolution Camera (HRC) for high-resolution investigations of rock texture at a distance. The cameras and electronics are housed in an optical bench that provides the mechanical interface to the rover mast and a planetary protection barrier. -

MULTI-SCALE ANALYSIS of ALLUVIAL SEDIMENTARY OUTCROPS in UTAH USING EXOMARS 2020 PANCAM, ISEM, and CLUPI EMULATORS
49th Lunar and Planetary Science Conference 2018 (LPI Contrib. No. 2083) 1911.pdf MULTI-SCALE ANALYSIS OF ALLUVIAL SEDIMENTARY OUTCROPS IN UTAH USING EXOMARS 2020 PANCAM, ISEM, and CLUPI EMULATORS. E. J. Allender1, C. R. Cousins1, M. D. Gunn2, R. Barnes3 1School of Earth and Environmental Sciences, University of St Andrews, Irvine Building, North St, St Andrews, UK ([email protected]), 2Department of Physics, Penglais Campus, Aberystwyth University, Aberystwyth, UK, 3Imperial College London, Kensington, London, UK. Introduction: As the launch date for the Instrument emulators were deployed at each site to ESA/Roscosmos ExoMars 2020 rover draws near, the simulate the PanCam, ISEM, and CLUPI instruments need for analysis tools to exploit the wealth of data to onboard ExoMars 2020. These were: the Aberstwyth be returned by its Panoramic Camera (PanCam) [1], University PanCam Emulator (AUPE) [8] (spanning Infrared Spectrometer for Mars (ISEM) [2], and 440-1000 nm), a Spectral Evolution spectrometer CLose-UP Imager (CLUPI) [3] instruments becomes (ISEM-E) spanning 440-2500 nm with hyperspectral increasingly important; the exploitation of integrated reolution, and CLUPI-E - a Sigma DSLR camera con- data from these instruments will be invaluable in de- figured to CLUPI specifications [3]. AUPE and tecting evidence of past habitability on Mars and iden- CLUPI-E mosaics were collected at each site, and tifying drilling locations. ISEM-E data points were collected either as vertical Together, PanCam, ISEM, and CLUPI offer multi- logs, or spanning each distinct unit. Soil and rock sam- scale analysis capabilities in both spatial (~140-1310 ples were also collected for each imaged unit at the µm/pixel at 2 m working distance), and spectral (350- study sites to provide spectral and mineralogical 3300 nm) dimensions. -

18Th EANA Conference European Astrobiology Network Association
18th EANA Conference European Astrobiology Network Association Abstract book 24-28 September 2018 Freie Universität Berlin, Germany Sponsors: Detectability of biosignatures in martian sedimentary systems A. H. Stevens1, A. McDonald2, and C. S. Cockell1 (1) UK Centre for Astrobiology, University of Edinburgh, UK ([email protected]) (2) Bioimaging Facility, School of Engineering, University of Edinburgh, UK Presentation: Tuesday 12:45-13:00 Session: Traces of life, biosignatures, life detection Abstract: Some of the most promising potential sampling sites for astrobiology are the numerous sedimentary areas on Mars such as those explored by MSL. As sedimentary systems have a high relative likelihood to have been habitable in the past and are known on Earth to preserve biosignatures well, the remains of martian sedimentary systems are an attractive target for exploration, for example by sample return caching rovers [1]. To learn how best to look for evidence of life in these environments, we must carefully understand their context. While recent measurements have raised the upper limit for organic carbon measured in martian sediments [2], our exploration to date shows no evidence for a terrestrial-like biosphere on Mars. We used an analogue of a martian mudstone (Y-Mars[3]) to investigate how best to look for biosignatures in martian sedimentary environments. The mudstone was inoculated with a relevant microbial community and cultured over several months under martian conditions to select for the most Mars-relevant microbes. We sequenced the microbial community over a number of transfers to try and understand what types microbes might be expected to exist in these environments and assess whether they might leave behind any specific biosignatures.