View of Evidence and Mechanisms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Aikawa Town Guide Book
Free Taking a look around Aikawa, So many things to do So many things to see! AikawaAikawa TownTown GuideGuide BookBook “Ai”“Ai” means means “Love”“Love”愛愛AiAi Aikawa is a town of love. So huge, so lovely! I’ m amazed Look, there are beautiful flowers at the power of the cascade! blooming throughout the year. (^o^) The beauty of this wonderful old residence shows the love of the carpenters of old. “450 years ago, a fierce battle took place and this stone marks the battleground.” I see... Town Areas and Sightseeing Spots Ken-O Expressway Wide Area Map Sagamihara IC Ken-O Expressway Sagamihara City Map of Places to See in AikawaKen-o-Do (Metropolitan Inter-City Expressway) Sagamihara City Aikawa Town Sagamihara Aikawa IC Horinouchi Uedana Kiyokawa Village Miyagase Dam: page 4 Hattori Dairy Farm: page 5 Mimase Park Atsugi City Battle of Mimase Pass: page 10 Mimase Park Athletic Ground Hattori Dairy Farm Rainbow Plaza Textile Association Prefectural Aikawa Park Shingen's Banner Tree Hinata Bridge Banda Station Hanbara Hinata Asari Grave Suzuki Confectionery and Shrine Miyagase Dam Aikawa Ohashi Bridge Boarding point for dam site pleasure boat Sagami Line Arts and Crafts Village Aikawa Forestry Association Mimase Prefectural Central Takata Bridge Water and Matsubazawa Fireflies Habitat High-tech Laboratory Estate Lake Miyagase Energy Museum Local museum Aikawa Bridge Suwa Shrine Mimase Takata Hashigiwa Osawa waterfall Prefectural Aikawa Hanbara Elem. Sagami river Ishigoya Dam Community Village Rankaya Hanbara Hanbara shogakko Iriguchi Sumida Kanagawa Central Poultry Farming Association Anzu no Shippo Bakery Battle of Mimase Pass Aikawa Solar Park battlefield marker Takamine Elem. -

Hydrochemistry of the Groundwaters in the Izu Collision Zone and Its Adjacent Eastern Area, Central Japan
Geochemical Journal, Vol. 45, pp. 309 to 321, 2011 Hydrochemistry of the groundwaters in the Izu collision zone and its adjacent eastern area, central Japan YOICHI MURAMATSU,1* YUTA NAKAMURA,2 JITSURO SASAKI3 and AMANE WASEDA4 1Department of Liberal Arts, Faculty of Science and Technology, Tokyo University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan 2Department of Chemistry, Faculty of Science, Tokyo University of Science, 1-3 Kagurazaka, Shinjuku-ku, Tokyo 162-8601, Japan 3Department of Pure and Industrial Chemistry, Faculty of Science and Technology, Tokyo University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan 4Japan Petroleum Exploration Co., Ltd., Research Center, 1-2-1 Hamada, Mihama-ku, Chiba 261-0025, Japan (Received April 8, 2010; Accepted March 6, 2011) Chemical and stable isotopic (δD, δ18O, δ34S) compositions of rivers and groundwaters, mineral constituents of rock samples from wells, and δ34S values of anhydrite in the Izu collision zone and its adjacent eastern area, southern Kanto Plain, central Japan, were analyzed to constrain the water-rock reactions and flow systems of the groundwaters. Inside the accreted Izu–Bonin–Mariana (IBM) basin, a two-dimensional map of the geothermal gradient calculated roughly using the discharge groundwater temperatures and the borehole temperature logging data confirms that the aqui- fer is recharged by the local meteoric water (LMW) and the high density seawater in the area. The oxygen and hydrogen isotopic compositions reveal that the Ca·Na–SO4 groundwaters in the Tanzawa Mts. and the high Na–Cl groundwaters in the coastal area are of meteoric water and weakly altered fossil seawater origins, respectively. -

Zootaxa 1325: 157–190 (2006) ISSN 1175-5326 (Print Edition) ZOOTAXA 1325 Copyright © 2006 Magnolia Press ISSN 1175-5334 (Online Edition)
Zootaxa 1325: 157–190 (2006) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA 1325 Copyright © 2006 Magnolia Press ISSN 1175-5334 (online edition) Geographic variation of chromosomes and somatic morphology in the Japanese polymorphic species Leiobunum hiraiwai (Arachnida: Opiliones: Sclerosomatidae) NOBUO TSURUSAKI Laboratory of Biology, Faculty of Regional Sciences, Tottori University, Tottori, 680-8551 Japan. E-mail: [email protected] Abstract The hiraiwai-subgroup of the Leiobunum curvipalpe-species group (Opiliones: Sclerosomatidae: Leiobuninae) is revised on the basis of somatic morphology, genitalia and karyotypes. This subgroup is distributed mainly in the mountainous areas of the Japanese Beech (Fagus crenata) forest zone (usually above 700 m in altitude in the southwestern part of Japan) in the western part of the Japanese Islands and corresponds to a single polytypic species, Leiobunum hiraiwai (Sato & Suzuki, 1939) with eight geographic races. Leiobunum tamanum Suzuki, 1957 and L. tsushimense Suzuki, 1976 are here synonymized with L. hiraiwai. All the races, except for the Tsushima race, were chromosomally analyzed. The number of chromosomes varies geographically from 2n = 18 to 22 eastwards, irrespective of the variation in somatic morphology or male genitalia. Component chromosomes in these karyotypes are invariably metacentric or submetacentric in structure. Some morphological characters show a geographic pattern of variation that conforms to the stasipatric model of speciation. The possible processes of geographical differentiation of the species and direction of character evolution are inferred on the basis of such geographical patterns. The distribution of some character states suggests that the closest relative of the subgroup is the curvipalpe-subgroup (= Leiobunum curvipalpe Roewer, 1910 and L. -

Mountaineering, Photographs, and World Heritage Sites Originally
Mountaineering, Photographs, and World Heritage Sites Originally told in Japanese by Hiroyuki Nakada Translated by Ryusui Seiryoin Cover Photo by Hiroyuki Nakada Cover design by Tanya Copyright © 2014 Hiroyuki Nakada / The BBB: Breakthrough Bandwagon Books All rights reserved. ISBN: 978-1-312-76695-2 The BBB website http://thebbb.net/ Hiroyuki Nakada Author Page http://thebbb.net/cast/hiroyuki-nakada.html 1. Starting Point of Mountaineering B (Editor of The BBB): Mr. Nakada, you are a mountaineering photographer and you hold the qualification of “World Heritage Meister,” the highest rank of the Test of World Heritage Study (which occurs only in Japan). First, we would like to ask you about mountaineering. To begin with, what was your first experience of mountains? What made you interested in mountains? Nakada: None of my family and acquaintances were interested in mountaineering, so I originally had nothing to do with outdoor life. I loved nature from my childhood, though. One thing comes to mind as a starting point. It was during my elementary school days. One summer day, on my way home from school, I saw an unbelievably gigantic cumulonimbus cloud as I was walking across a pedestrian overpass. The beauty of the contrast between the huge white cloud and clear blue sky fascinated me. I came to feel as if the cloud were a snow mountain. At the time, I imagined that if I could climb such a great mountain, it would be the best experience. I still remember the scene vividly. I think it was the very moment when I first became interested in mountains. -

Energy Efficiency and GHG Reduction Final Report
Public Disclosure Authorized Study of Japanese Experiences Public Disclosure Authorized on Sustainable Urban Development including Pollution Control and Management, Resource/Energy Efficiency and GHG Reduction Public Disclosure Authorized FINAL REPORT February 2011 Public Disclosure Authorized THE WORLD BANK JAPAN INTERNATIONAL COOPERATION AGENCY STUDY OF JAPANESE EXPERIENCES ON SUSTAINABLE URBAN DEVELOPMENT INCLUDING POLLUTION CONTROL AND MANAGEMENT, RESOURCE / ENERGY EFFICIENCY AND GHG REDUCTION FINAL REPORT The First East Asia Eco2 Program, including this study, was funded by the Cities Alliance through a non-core contribution of the Japanese Government, Japan International Cooperation Agency (JICA), and the World Bank February 2011 ALMEC CORPORATION TABLE OF CONTENTS MAIN TEXT 1 SUMMARY 1.1 Context of the Study.......................................................................................................1-1 1.2 Study Objectives............................................................................................................1-2 1.3 Analytical Framework of the Eco2 Initiative....................................................................1-3 1.4 Urban Development Process, Urban Management, and Environmental Initiatives in Japan..........................................................................................................................1-5 1.5 Responses of Stakeholders...........................................................................................1-8 1.6 Lessons from Japanese Experiences..........................................................................1-11 -
Open to the Public
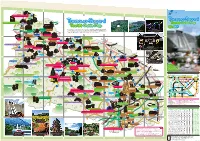
A B C D E F 521 Wada no Sato Experience Center "Village House" G H Mt. Jimba Tanzawa-Sagami 100 Japanese Rural Villages・ Sanogawa Area To Hachioji Tanzawa Sagami Jimba no Yu - Tourist Guide Map 522 1 Touristst GuGuiddee Mapp 1 たび相模 Ishii House 旅 丹沢・相模 観光ナビ Expresswa uo y Ch Sagamiko-Higashi Kanagawa Prefectural Government Website: Tanzawa-Sagami Tourist Guide To Kofu Obarajuku Honjin Midori's Love Letter Lake Tsukui Ogura Bridge Sagamiko IC IC provides sightseeing information: places to visit, souvenirs to purchase, and Midori Ward Sagamihara City (Fujino area) Midori Ward Sagamihara City (Tsukui area) Midori Ward Sagamihara City (Shiroyama area) K MAP - MAP - MAP - osh A 2 /D 2 Guide: Nature /E 2 Guide: History u Higway e in . It's updated regularly. Check it out online! L 20 To Takao events to experience 20 uo Ch Fujino JR Sagamiko Lake Sagami Yoshino-juku Fujiya Sagamiko Memorial Hall Prefectural Sagamiko Park To Hachioji Lake Shiroyama Shiroyama lake promenade To Hachioji To Hachioji Art Walk Sagamiko Resort Pleasure Forest Shokakuji Temple 515 Sagamihara City Mt.Sekiro To Chofu Ozaki Gakudo Memorial House Midori Ward Lake Tsukui 413 (Shiroyama Area) Hashimoto e Fujino Hot Springs Higashiotaru no Yu Lin ihara gam io Sa 2 Prefectural Fujino Art House Sagamihara City 412 Sagamihara City Ke 2 517 J Midori Ward Midori Ward R Y ok Obarajuku Honjin Shinohara no Sato Center oh (Sagamiko Area) am a Midori Ward Sagamihara City (Sagamiko area) Lin Ogura Bridge e /MAP C-1 Guide: History Fujino Yamanami Hot Springs IC Sagamihara Sagamigawa Nature Village Park 518 Sagamihara Prefectural Tsukuiko-Shiroyama Park 508 Minami- 129 Hashimoto Sagamihara City Yabe Kami-Ooshima Campgrounds Midori Ward 16 Doshi Riv. -

Modeling of Species Geographic Distribution for Assessing Present Needs for the Ecological Networks
MODELING OF SPECIES GEOGRAPHIC DISTRIBUTION FOR ASSESSING PRESENT NEEDS FOR THE ECOLOGICAL NETWORKS T. Dokoa, *,F A. Kooimanb, A.G.Toxopeusb aGraduate School of Media and Governance, Keio University, 302 Z-building of Keio University, 5322 Endoh, Fujisawa, Kanagawa, 252-8520, Japan - [email protected] bDepartment of Natural Resources, International Institute for Geo-information Science and Earth Observation (ITC), P. O. Box 6, Hengelosestraat 99, 7500 AA, Enschede, The Netherlands - [email protected], [email protected] KEY WORDS: Ecology, Environment, GIS, Modelling, Algorithms, Landscape, Method, Proposal ABSTRACT: In Japan, attention is currently focused on designing ecological networks for wildlife animals. However there is an obvious lack of the species spatial information. This study aims (a) to acquire the potential spatial distribution of Asiatic black bear and Japanese se- row to identify core areas, and (b) to propose a methodology for assessing needs for ecological networks. 1836 species’ point records and 14 potential predictors were prepared in a GIS environment, split into a train and a test dataset. Screening predictors by statisti- cal analysis, we modeled species geographic distribution by three algorithms: GARP, MaxEnt, and GLMs in Kanagawa and Shizu- oka Prefectures. Based on the most accurate maps, assessed by Kappa statistics, population was estimated based on population den- sity and area of habitat patch. For bear, MaxEnt performed best with the predictor variables: altitude, distance to paths and stone steps, distance to wide roads, and vegetation cover types. GARP failed to predict presence in Fuji. Its best GLM equation was log(p/(1-p))=(-1.486e+01)+(7.335e-04)*distance to paths and stone steps+(9.470e-03)*altitude. -

Climbing a Few of Japan's 100 Famous Mountains – Volume 4: Mt
COPYRIGHTED MATERIAL Climbing a Few of Japan’s 100 Famous Mountains – Volume 4: Mt. Hakkoda & Mt. Zao Daniel H. Wieczorek and Kazuya Numazawa COPYRIGHTEDMATERIAL Climbing a Few of Japan’s 100 Famous Mountains – Volume 4: Mt. Hakkoda & Mt. Zao COPYRIGHTEDMATERIAL Copyright © 2014 Daniel H. Wieczorek and Kazuya Numazawa All rights reserved. ISBN-10: 0996216162 ISBN-13: 978-0-9962161-6-6 DEDICATION This work is dedicated, first of all, to my partner, Kazuya Numa- zawa. He always keeps my interest in photography up and makes me keep striving for the perfect photo. He also often makes me think of the expression “when the going gets tough, the tough keep going.” Without my partner it has to also be noted that I most likely would not have climbed any of these mountains. Secondly, it is dedicated to my mother and father, bless them, for tolerating and even encouraging my photography hobby from the time I was twelve years old. And, finally, it is dedicated to my friends who have encouraged me to create books of photographs which I have taken while doing mountain climbing. COPYRIGHTED MATERIAL Other Books in this Series “Climbing a Few of Japan's 100 Famous Mountains – Volume 1: Mt. Daisetsu (Mt. Asahidake)”; ISBN-13: 9781493777204; 66 Pages; Dec. 5, 2013 “Climbing a Few of Japan's 100 Famous Mountains – Volume 2: Mt. Chokai (Choukai)”; ISBN-13: 9781494368401; 72 Pages; Dec. 8, 2013 “Climbing a Few of Japan's 100 Famous Mountains – Volume 3: Mt. Gassan”; ISBN-13: 9781494872175; 70 Pages; Jan. 4, 2014 “Climbing a Few of Japan's 100 Famous Mountains – Volume 5: Mt. -

(PDF): SFCJ9-01.Pdf
Geospatial Approaches towards Biodiversity Conservation ◆特集*招待論文◆ Geospatial Approaches towards Biodiversity Conservation Case study in Fuji-Tanzawa regions in Japan 生物多様性保全に向けた 空間情報学的アプローチ 富士丹沢地域を事例研究として Tomoko Doko Doctoral Program, Graduate School of Media and Governance, Keio University Research Fellow of the Japan Society for the Promotion of Science 土光 智子 慶應義塾大学大学院政策・メディア研究科後期博士課程 日本学術振興会特別研究員 (DC) Hiromichi Fukui 福井 弘道 Professor, Faculty of Policy Management, Keio University 慶應義塾大学総合政策学部教授 In this paper, we introduce several geospatial approaches for biodiversity conservation, taking an example of our on-going project. Chapter 1 introduces the needs of ecological networks formation, and Fuji-Tanzawa regions as the study area. Chapter 2 presents the ecological modeling approach which can model species’ geographic distribution and assess present needs for the ecological networks. Chapter 3 outlines fieldwork for Asiatic black bears’ ecology, which focuses on the capture and collecting fieldsigns. Chapter 4 introduces few experimental results on an advanced satellite tracking system, namely GPS-AROGS, concerning animal movement. Chapter 5 presents a Web-based GIS development as a policy making support system for conserving biodiversity. Chapter 6 briefly described future works. 本稿では、生物多様性保全に向けた空間情報アプローチを紹介する。第 1 章では、エコロジカル ネットワークと研究対象地域の富士丹沢地域を概観した。第 2 章では、生息地域空間的分布モデリ ングを用いて、エコロジカルネットワークの必要性をアセスメントする定量的手法を提案した。第 3 章は、ツキノワグマの生態を調査するフィールドワーク方法を紹介した。第 4 章では、最新型の 動物衛星追跡システムの精度実証実験を紹介した。第 5 章においては、政策決定支援システムとし て、Web-GIS を開発した。第 6 章では、将来展望を述べた。 Keywords: Ecological networks, Asiatic black bear, ARGOS, Modeling, Web-GIS KEIO SFC JOURNAL Vol.9 No.1 2009 特集 位置情報計測技術の応用 1 Introduction belt. Vegetation is varied from the plants in the warm- 1.1 Biodiversity loss and ecological networks temperature zone in Izu Island chain to the ones in International awareness of the need to protect the Alperstein zone of Mt. -

Mt. Asahidake)
Climbing a Few of Japan’s 100 Famous Mountains – Volume 1: Mt. Daisetsu (Mt. Asahidake) Daniel H. Wieczorek and Kazuya Numazawa COPYRIGHTED MATERIAL Climbing a Few of Japan’s 100 Famous Mountains – Volume 1: Mt. Daisetsu (Mt. Asahidake) COPYRIGHTEDMATERIAL Copyright © 2014 Daniel H. Wieczorek and Kazuya Numazawa All rights reserved. ISBN: 1493777203 ISBN-13: 978-1493777204 DEDICATION This work is dedicated, first of all, to my Secondly, it is dedicated to my mother and partner, Kazuya Numazawa. He always keeps father, bless them, for tolerating and even my interest in photography up and makes me encouraging my photography hobby from the keep striving for the perfect photo. He also time I was 12 years old. often makes me think of the expression “when the going gets tough, the tough keep going.” And, finally, it is dedicated to my friends who Without my partner it has to also be noted have encouraged me to create books of that I most likely would not have climbed any photographs which I have taken while doing of these mountains. mountain climbing. COPYRIGHTEDMATERIAL Other books by Daniel H. Wieczorek and Kazuya Numazawa “Outdoor Photography of Japan: Through the Seasons”; ISBN/EAN13: 146110520X / 9781461105206; 362 Pages; June 10, 2011; Also available as a Kindle Edition “Some Violets of Eastern Japan”; ISBN/EAN13: 1463767684 / 9781463767686; 104 Pages; August 20, 2011; Also available as a Kindle Edition “2014 Photo Calendar – Japan's Flowers, Plants & Trees”; ISBN/EAN13: 1482315203 / 978- 1482315202; 30 Pages; February 4, 2013 “2014 Photo Calendar – Japan Mountain Scenery”; ISBN/EAN13: 1482371383 / 978- 1482371383; 30 Pages; February 15, 2013 Other books by Daniel H. -

Mt. Shirane (Kusatsu)
Climbing a Few of Japan’s 100 Famous Mountains – Volume 6: Mt. Shirane (Kusatsu) Daniel H. Wieczorek and Kazuya Numazawa COPYRIGHTED MATERIAL Climbing a Few of Japan’s 100 Famous Mountains – Volume 6: Mt. Shirane (Kusatsu) COPYRIGHTEDMATERIAL Copyright © 2014 Daniel H. Wieczorek and Kazuya Numazawa All rights reserved. ISBN: 1497303230 ISBN-13: 978-1497303232 DEDICATION This work is dedicated, first of all, to my Secondly, it is dedicated to my mother and partner, Kazuya Numazawa. He always keeps father, bless them, for tolerating and even my interest in photography up and makes me encouraging my photography hobby from the keep striving for the perfect photo. He also time I was 12 years old. often makes me think of the expression “when the going gets tough, the tough keep going.” And, finally, it is dedicated to my friends who Without my partner it has to also be noted have encouraged me to create books of that I most likely would not have climbed any photographs which I have taken while doing of these mountains. mountain climbing. COPYRIGHTEDMATERIAL Other books by Daniel H. Wieczorek and Kazuya Numazawa “Outdoor Photography of Japan: Through the Seasons”; ISBN/EAN13: 146110520X / 9781461105206; 362 Pages; June 10, 2011; Also available as a Kindle Edition “Some Violets of Eastern Japan”; ISBN/EAN13: 1463767684 / 9781463767686; 104 Pages; August 20, 2011; Also available as a Kindle Edition “2014 Photo Calendar – Japan's Flowers, Plants & Trees”; ISBN/EAN13: 1482315203 / 978- 1482315202; 30 Pages; February 4, 2013 “2014 Photo Calendar – Japan Mountain Scenery”; ISBN/EAN13: 1482371383 / 978- 1482371383; 30 Pages; February 15, 2013 “Climbing a Few of Japan's 100 Famous Mountains – Volume 1: Mt. -

Night & Morning
NEWS TOPICS Aug. 8, A next-generation entertainment Eat, play and experience at an Mar. 15, ome to Kanaga 2019 2019 elc wa NEW restaurant based on a ukiyo-e theme! entertainment building complex NEW W OPEN UKIYO-E CAFÉ ASOBUILD OPEN https://www.ukiyoecafe.com/?lang=en https://asobuild.com/en/ 17:00- Enjoy a show that fuses digital Try out a variety of activities in this complex 18:00- 21:30 projections with non-verbal including a floor at which you can participate 22:00 performances by the world-class in workshops such as a ceramic art workshop, Night & Morning performance troupe SIRO-A. Within Ei and a restaurant floor. 2-14-9 Takashima, Ichibankan, 6F Kanagawa Kenmin Hall, Nishi-ku, Yokohama City 10:00-22:00 3-1 Yamashita-cho, Naka-ku, Yokohama (Vary by floor and shop) Irregular holidays City 2 performances from 18:00 and Directly linked to the South East Exit amazing spots!! from 20:00 Tues. & Wed. Approx. passage of Yokohama Station (Several lines, 6 min. walk from Minatomirai Line Nihon-odori Sta. (MM 05) Station No.: KK 37/TY 21/MM 01, etc.) INFORMATION South of Tokyo,Japan ACCESS to KANAGAWA Tokaido Main Line, about 10 min. Narita Express Kawasaki Limited Express, Shinagawa JR Tokaido Shinkansen, Hakone Tozan Line, about 70 min. about 30 min. about 15 min. Odawara Hakone Yumoto Narita Minatomirai Line, about 3 min. Narita Express Minatomirai Airport Limited Express, Yokosuka Line, about 90 min. Yokosuka Line, about 18 min. about 6 min. Yokohama Ofuna Kamakura Keikyu Line Limited Express, about 45 min.