Further Observations on Use of the Feet by Foraging Herons
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Florida Keys Vessel Pumpout Facilities Marine Sanitation Device
Marine Sanitation Device Discharge Regulations Effective: December 27, 2010 Activities prohibited Sanctuary-Wide: q Discharge of sewage incidental to vessel use and generated by a marine sanitation device in accordance with the Federal Water Pollution Control Act (also called the Clean Water Act). q Having a marine sanitation device that is not secured in a manner that prevents discharges or deposits of treated and untreated sewage. Acceptable methods include, but are not limited to, all methods that have been approved by the U.S. Coast Guard. Pumpout facilities are located throughout the Keys to assist boat operators in complying with this rule. For a list of pumpout facilities, visit http://www.dep.state.fl.us/cleanmarina/about.htm. Florida Keys Vessel Pumpout Facilities * Designated Clean Marina Facility Key West Duck Key Mobile Pumpout Services • A & B Marina • Hawk’s Cay Resort Marina Free pumpout services for vessels • Conch Harbor Marina* anchored within unincorporated Long Key • City Marina at Garrison Bight* Monroe County (Key Largo, • Key West Bight Marina* • Fiesta Key KOA Tavernier, Cudjoe, Big Pine, Stock Stock Island Upper Matecumbe Key Island, etc.) and the Village of • Stock Island Marina Village • Bayside Marina- World Wide Sportsman* Islamorada. • Sunset Marina • Coral Bay Marina • Pumpout USA at 305-900-0263 or visit www.po-keys.com. Lower Keys Plantation Key • Plantation Yacht Harbor* • Bahia Honda State Park* • City of Key West 305-292-8167 • Sunshine Key RV Resort & Marina • Treasure Harbor Marine* • Stock Island, Mark LPS 305-587-2787 Marathon Tavernier • City of Marathon 305-289-8877 • Boot Key Harbor City Marina • Mangrove Marina • Key Colony Beach 305-289-1310 • Burdines Waterfront • Marathon Yacht Club Key Largo • Panchos Fuel Dock & Marina • All Keys Portalet Tips: • Sombrero Marina Dockside* • John Pennekamp Coral Reef State Park* • Check with marina ahead of time on • Manatee Bay Marina status of pumpout equipment. -
Key West Stock Island Key Haven
Key Haven Stock Island 7 Key West 11 9 1 )"5 )"3 )"4 11 1 11 10 9 11 7 )"2 5 2 1 )"0 )" 8 6 4 3 Monroe County - Voting Precincts : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' Data Source: Monroe County - Florida - Supervisor of Elections Date: 1/30/2019 12 )"12 Big Coppitt Key )"11 Boca Chica Key 12 )"10 )"9 )"8 12 12 13 )"7 )"6 12 12 Geiger Key 12 Monroe County - Voting Precincts : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' Data Source: Monroe County - Florida - Supervisor of Elections Date: 1/30/2019 )"16 13 13 13 13 Sugarloaf Key Saddlebunch Keys )"15 13 13 13 13 13 )"14 13 13 )"13 13 13 13 13 )"12 13 13 Monroe County - Voting Precincts : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' Data Source: Monroe County - Florida - Supervisor of Elections Date: 1/30/2019 13 13 Sugarloaf Key 13 )"19 13 13 13 13 )"18 13 )"17 Monroe County - Voting Precincts 13 : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' Data Source: Monroe County - Florida - Supervisor of Elections 13 Date: 1/30/2019 Cudjoe Key 14 15 )"21 )"22 Summerland Key )"23 )"24 15 )"25 )"20 )" 13 15 Sugarloaf Key 13 13 Monroe County - Voting Precincts 13 : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' Data Source: Monroe County - Florida - Supervisor of Elections Date: 1/30/2019 Big Torch Key 15 Cudjoe Key 14 Monroe County - Voting Precincts 15 : )" US1 Mile Markers Voting Precinct Boundary 1 " = 2,000 ' 13Data Source: Monroe County - Florida - Supervisor of Elections Date: 1/30/2019 Big Pine Key 30 15 Little Torch -

Monroe County Stormwater Management Master Plan
Monroe County Monroe County Stormwater Management Master Plan Prepared for Monroe County by Camp Dresser & McKee, Inc. August 2001 file:///F|/GSG/PDF Files/Stormwater/SMMPCover.htm [12/31/2001 3:10:29 PM] Monroe County Stormwater Management Master Plan Acknowledgements Monroe County Commissioners Dixie Spehar (District 1) George Neugent, Mayor (District 2) Charles "Sonny" McCoy (District 3) Nora Williams, Mayor Pro Tem (District 4) Murray Nelson (District 5) Monroe County Staff Tim McGarry, Director, Growth Management Division George Garrett, Director, Marine Resources Department Dave Koppel, Director, Engineering Department Stormwater Technical Advisory Committee Richard Alleman, Planning Department, South Florida WMD Paul Linton, Planning Department, South Florida WMD Murray Miller, Planning Department, South Florida WMD Dave Fernandez, Director of Utilities, City of Key West Roland Flowers, City of Key West Richard Harvey, South Florida Office U.S. Environmental Protection Agency Ann Lazar, Department of Community Affairs Erik Orsak, Environmental Contaminants, U.S. Fish and Wildlife Service Gus Rios, Dept. of Environmental Protection Debbie Peterson, Planning Department, U.S. Army Corps of Engineers Teresa Tinker, Office of Planning and Budgeting, Executive Office of the Governor Eric Livingston, Bureau Chief, Watershed Mgmt, Dept. of Environmental Protection AB i C:\Documents and Settings\mcclellandsi\My Documents\Projects\SIM Projects\Monroe County SMMP\Volume 1 Data & Objectives Report\Task I Report\Acknowledgements.doc Monroe County Stormwater Management Master Plan Stormwater Technical Advisory Committee (continued) Charles Baldwin, Islamorada, Village of Islands Greg Tindle, Islamorada, Village of Islands Zulie Williams, Islamorada, Village of Islands Ricardo Salazar, Department of Transportation Cathy Owen, Dept. of Transportation Bill Botten, Mayor, Key Colony Beach Carlos de Rojas, Regulation Department, South Florida WMD Tony Waterhouse, Regulation Department, South Florida WMD Robert Brock, Everglades National Park, S. -

Mile Marker 0-65 (Lower Keys)
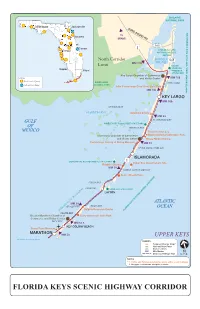
Key to Map: Map is not to scale Existing Florida Keys Overseas Heritage Trail Aquatic Preserves or Alternate Path Overseas Paddling Trail U.S. 1 Point of Interest U.S. Highway 1 TO MIAMI Kayak/Canoe Launch Site CARD SOUND RD Additional Paths and Lanes TO N KEY LARGO Chamber of Commerce (Future) Trailhead or Rest Area Information Center Key Largo Dagny Johnson Trailhead Mangroves Key Largo Hammock Historic Bridge-Fishing Botanical State Park Islands Historic Bridge Garden Cove MM Mile Marker Rattlesnake Key MM 105 Florida Department of Environmental Protection, Office of Greenways & Trails Florida Keys Overseas Heritage Trail Office: (305) 853-3571 Key Largo Adams Waterway FloridaGreenwaysAndTrails.com El Radabob Key John Pennekamp Coral Reef State Park MM 100 Swash Friendship Park Keys Key Largo Community Park Florida Keys Community of Key Largo FLORIDA BAY MM 95 Rodriguez Key Sunset Park Dove Key Overseas Heritage Trail Town of Tavernier Harry Harris Park Burton Drive/Bicycle Lane MM 90 Tavernier Key Plantation Key Tavernier Creek Lignumvitae Key Aquatic Preserve Founders Park ATLANTIC OCEAN Windley Key Fossil Reef Geological State Park MM 85 Snake Creek Long Key Historic Bridge TO UPPER Islamorada, Village of Islands Whale Harbor Channel GULF OF MEXICO KEYS Tom's Harbor Cut Historic Bridge Wayside Rest Area Upper Matecumbe Key Tom's Harbor Channel Historic Bridge MM 80 Dolphin Research Center Lignumvitae Key Botanical State Park Tea Table Key Relief Channel Grassy Key MM 60 Conch Keys Tea Table Channel Grassy Key Rest Area Indian Key -

03.01.2012 WKFRGSP AP.Pdf
Windley Key Fossil Reef Geological State Park APPROVED Unit Management Plan STATE OF FLORIDA Department of Environmental Protection Division of Recreation and Parks March 1, 2012 TABLE OF CONTENTS INTRODUCTION .............................................................................................................1 PURPOSE AND SIGNIFICANCE OF THE PARK .....................................................1 PURPOSE AND SCOPE OF THE PLAN ......................................................................2 MANAGEMENT PROGRAM OVERVIEW ................................................................8 Management Authority and Responsibility ..............................................................8 Park Management Goals ..............................................................................................9 Management Coordination ........................................................................................10 Public Participation .....................................................................................................10 Other Designations ......................................................................................................10 RESOURCE MANAGEMENT COMPONENT INTRODUCTION ...........................................................................................................11 RESOURCE DESCRIPTION AND ASSESSMENT .................................................12 Natural Resources .......................................................................................................12 Topography -

Florida Keys Overseas Heritage Trail
9/15 FloridaStateParks.org/floridakeys FloridaGreenwaysAndTrails.com. (305) 853-3571 (305) 3 La Croix Court, Key Largo, FL 33037 FL Largo, Key Court, Croix La 3 of the Florida Keys Overseas Paddling Trail, visit Trail, Paddling Overseas Keys Florida the of Florida Keys Overseas Heritage Trail Heritage Overseas Keys Florida of Greenways & Trails. For maps and descriptions and maps For Trails. & Greenways of Division of Recreation and Parks and Recreation of Division Paddling Trail, a project coordinated by the Office the by coordinated project a Trail, Paddling Florida Department of Environmental Protection Environmental of Department Florida 1500-mile Florida Circumnavigational Saltwater Circumnavigational Florida 1500-mile the environment. The Trail is a segment of the of segment a is Trail The environment. the recreational opportunities that minimally impact minimally that opportunities recreational and ecology of the Keys while providing while Keys the of ecology and The “blueway” introduces visitors to the history the to visitors introduces “blueway” The Overseas Heritage Trail. Trail. Heritage Overseas the growing sport of kayak and canoe touring. canoe and kayak of sport growing the of Recreation and Parks as part of the Florida Keys Florida the of part as Parks and Recreation of a paddling destination for those involved in involved those for destination paddling a Department of Environmental Protection’s Division Protection’s Environmental of Department The Florida Keys Overseas Paddling Trail offers Trail Paddling Overseas Keys Florida The Historic Places and are managed by the Florida the by managed are and Places Historic have been listed on the National Register of Register National the on listed been have Trail all of the remaining 23 Flagler Railroad Bridges Railroad Flagler 23 remaining the of all Florida Keys Overseas Paddling Paddling Overseas Keys Florida into an economically viable destination. -

Welcome to Islamorada, Village of Islands, Florida
— POSITION AVAILABLE — VILLAGE MANAGER | Apply by April 9, 2021 Welcome to Islamorada, Village of Islands, Florida If you are looking for water, sun, expansive ocean and bay views, great restaurants, and a small-town atmosphere, read on. Islamorada is the jewel of the Florida Keys, and indeed all of Florida. Split lengthwise by the scenic Overseas Highway, it stretches 18 linear miles and is composed of four main islands�Plantation Key, Windley Key, Upper Matecumbe Key and Lower Matecumbe Key. Clear night skies and low levels of light pollution make visibility of the stars and planets one of the area’s many attractions. If you and your family love the water and the outdoors, Islamorada is paradise. Nestled between the Atlantic Ocean and the Gulf of Mexico, Islamorada is a great place for a fisherman. Tarpon, grouper, hogfish, snook, snapper, mahi mahi, sailfish, and permit are all plentiful here. That is why Islamorada is known as the Sport Fishing Capital of the World. Don’t have a boat? Don’t worry. CITY MANAGER | v CRESCENT CITY, FLORIDA 1 Kayaks and charters are readily available as are offshore and nurse sharks. Dolphins put on quite a show swimming, jumping backcountry fishing guides. Or just throw your line in from one through a hoop and ringing a bell. The 17-acre site has exotic ofthebridges. Ifyouwouldratherwatchthefishinstead,thearea birds, lizards, crocodilians, sea turtles, tropical game fish, sharks, is known for its snorkeling and scuba diving. You can visit reefs andotherformsofmarinelife.Shortcruisesandbottomlessboat and shipwrecks, or just prowl the bottom. Do you like lobster? rides (that is, a boat with an open center) are offered here. -
FL Keys Application Map 11X17.Ai
BISCAYNE NATIONAL PARK Tallahassee Jacksonville CARD SOUND RD To Daytona MIAMI Orlando 905 Cocoa 1 Tampa CROCODILE LAKE NATIONAL WILDLIFE REFUGE North Corridor BARNES 905 MM 110 SOUND Limit DAGNY Naples JOHNSON Miami HAMMOCK STATE PARK Key Largo Chamber of Commerce LEGEND and Visitor Center MM 106 Florida Scenic Highway EVERGLADES KEY LARGO National Scenic Byway NATIONAL PARK John Pennekamp Coral Reef State Park MM 102 JOHN PENNEKAMP CORAL REEF STATE PARK (UNDERWATER) KEY LARGO MM 100 OYSTER KEYS FLORIDA BAY Wild Bird Center MM 93 GULF PLANTATION KEY WINDLEY KEY FOSSIL REEF STATE PARK OF WINDLEY KEY MEXICO Theater of the Sea Islamorada Chamber of Commerce Marine Mammal Adventure Park and Visitor Center Whale Harbor Marina Florida Keys History of Diving Museum MM 83 UPPER MATECUMBE KEY ISLAMORADA LIGNUMVITAE KEY BOTANICAL STATE PARK 1 Indian Key State Historic Site Robbie’s Marina MM 78 LOWER MATECUMBE KEY Anne’s Beach Park ARY FIESTA KEY LONG KEY LONG KEY STATE PARK LAYTON ATLANTIC MM 59 GRASSY KEY DUCK KEY OCEAN Dolphin Research Center INTRACOASTAL WATERWAY FLORIDA KEYS NATIONAL MARINE SANCTU CRAWL KEY Greater Marathon Chamber of Curry Hammock State Park Commerce and Visitor Center VACA KEY MM 53.5 KEY COLONY BEACH Crane Point Museum MARATHON MM 50 UPPER KEYS continued on next page LEGEND xxx Features/Itinerary Stops* xxx National/State Parks xxx Visitor Centers MM Mile Marker Overseas Heritage Trail NOTES * 1. Diving and Fishing opportunities along entire scenic highway. 2. No gaps or intrusions along the corridor. FLORIDA KEYS SCENIC -

Archaeological Investigations on the Upper Florida Keys by JOHN M
Archaeological Investigations On The Upper Florida Keys by JOHN M. GOGGIN D URING THE SUMMER of 1940 an archaeological survey was made of a portion of the Upper Florida Keys.' The area covered was from Virginia and Biscayne Keys on the north to Lower Mate- cumbe Key on the south, plus a portion of the west shore of Biscayne Bay. Summer is the least desirable time to attempt work in this area, but circumstances offered no other choice. The area in question was inhabited by the Tekesta* Indians.2 Ap- parently this tribe, like various others in the state, was more or less a political confederation of small local groups. There are many references to the presence of Indians on the Keys or "Los Martires" as the Spanish frequently called them, but few villages are named. Fontaneda does mention some towns, but they appear to have been located on the Lower Keys. In 1607 Governor Ibarra states that he received a visit from some chiefs, among whom were " . .. principal lords of the mouth of Miguel Mora." According to Swanton ".. this name was given to the opening between the Florida Mainland and the Keys on the eastern side." 3 This may be the present Card Sound. The one tribe or village that is men- tioned rather commonly is Matecumbe. This was probably located on the present Lower Matecumbe Key, because on many early maps it is called Old Matecumbe to differentiate it from Upper Matecumbe Key. The earliest mention is in 1573, when the Matecumbe Indians attained notice by killing a number of Spaniards.4 It was again mentioned about 1. -

Gulf of Mexico Bay Florida a Di ATLANTICOCEAN L F
Flo Shark 94 Key Big Cypress rid Everglades National Valley Biscayne National Preserve a 0 20 km 's 0 10 miles Turnpike 997 Peters Boca (to Biscayne Everglades Goulds Chita Gulf of ll) P National ark B National Park Key Mexico Park oun 821 Biscayne Florida d Homestead a Bay Keys ry Florida City National 9336 Adams T h e E v e r g l a d e s Elliott Marine C Key Key a Sanctuary rd So INSET Shark Point u n d Rd Card Middle Sound Hospital Whitewater Key Bay Key East Barnes John Pennekamp Loggerhead Key Sound 905 Key Cape Coral Reef Bush Key Florida Keys Marquesas Sable Key State Park Garden Key Long Key National Marine Keys Largo Sanctuary Dry Tortugas National Park Key Largo Flamingo Key National Key West National Largo Marine Wildlife Refuge Sanctuary Note: Same scale as main map Florida Bay Tavernier Plantation Key Windley Key Plantation Upper Matecumbe Key Shell Key Islamorada Lignumvitae Robbie's Marina Key Indian Key State Historic Site s Lower y Great White e Heron Layton Long Matecumbe K See Inset National Key Key r Little Wildlife Conch Key 1 p e Marathon Long Key State p Torch Refuge Duck Key U Big Torch Key Airport Recreation Area Little Pine Grassy Key Great White Key a Heron Big Pine Key Mile Marathon Florida Keys Florida Keys Seven i d National Marine National Cudjoe Key No Crane Point Museum National Marine r Snipe Key Bridge Sanctuary o Sanctuary Wildlife Keys Name Boot Key Vaca s l Refuge Key PigeonKey e y F Big Bahia Key K f Marquesas Sugarloaf Pine Honda l e o Key West Boca Chica d s Keys 1 Key Key i d i t National Key Summerland Ramrod Bahia M r a A T L A N T I C Wildlife Key t Key West Key Honda S Refuge State Park O C E A N Key West Key International Looe Key West Airport National Marine Sanctuary y s K e e r L o w. -

Florida Keys Overseas Heritage Trail Includes Includes Trail Heritage Overseas Keys Florida He T
PO00306_Rev_03_2020 bicyclists and other trail users. trail other and bicyclists historic bridges are popular with anglers, hikers, hikers, anglers, with popular are bridges historic Cut bridge and Tom’s Harbor Channel bridge. The The bridge. Channel Harbor Tom’s and bridge Cut FLStateParks # from the Long Key historic bridge, Tom’s Harbor Harbor Tom’s bridge, historic Key Long the from s on social media social on s u Follow watchers. Farther down the trail, enjoy the views views the enjoy trail, the down Farther watchers. FloridaStateParks.org stop at Long Key State Park, a popular spot for bird- for spot popular a Park, State Key Long at stop Channel 2 and Channel 5 bridges. Make another another Make bridges. 5 Channel and 2 Channel opular fishing spots in the area include the historic historic the include area the in spots fishing opular P Overseas Heritage Trail. Heritage Overseas Archaeological Preserve State Park. State Preserve Archaeological of Recreation and Parks as part of the Florida Keys Keys Florida the of part as Parks and Recreation of Historic State Park and San Pedro Underwater Underwater Pedro San and Park State Historic Department of Environmental Protection’s Division Division Protection’s Environmental of Department Lignumvitae Key Botanical State Park, Indian Key Key Indian Park, State Botanical Key Lignumvitae of Historic Places and are managed by the Florida Florida the by managed are and Places Historic of the Florida Keys. Side trips on the water can include include can water the on trips Side Keys. Florida the railroad bridges are listed on the National Register Register National the on listed are bridges railroad Geological State Park to learn about the geology of of geology the about learn to Park State Geological recreation, and historical reflection. -

Florida Keys Sea Heritage Journal
$2 Florida Keys Sea Heritage Journal VOL. 16 NO. 4 SUMMER 2006 USS SHARK OFFICIAL QUARTERLY PUBLICATION OF THE KEY WEST MARITIME HISTORICAL SOCIETY Labor Day: Night Of Harrow Mozelle Williams’ Handwritten Account of the 1935 Labor Day Hurricane Commentary and Transcription By Thomas Neil Knowles Copyright 2006 On the evening of September 2, 1935, the first Category V hurricane to make landfall in the United States cut a swath of death and destruction across the Florida Keys. The center crossed the Florida East Coast Railway track at Craig with maximum winds estimated by the Weather Bureau at over 200 mph. According to the U.S. Army Corps of Engineers, the water surged across the low-lying islands from Long Key to the south end of Key Largo reaching a maximum height of 18 feet above mean sea level. The hurricane originated east of the Bahamas and had been tracked for several On Upper Matecumbe Key where days by the Weather Bureau’s hurricane center in Jacksonville, Florida. A compact the most powerful part of the storm weather system that was difficult to precisely locate with the technology available came ashore, two families had taken in 1935, the eye of the hurricane had been projected to pass south of Key West, but shelter inside a heavily braced instead passed over Craig near the middle of the Florida Keys. Photo credit: the storm house. When that began to Author come apart, the four adults and five girls (9 & 8 years old) and a boy (18 raised in the Keys, Eddie had a boat children tried to jam themselves months).