Wyoming Animal Damage Management Board 2005 Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Page 5 of the 2020 Antelope, Deer and Elk Regulations
WYOMING GAME AND FISH COMMISSION Antelope, 2020 Deer and Elk Hunting Regulations Don't forget your conservation stamp Hunters and anglers must purchase a conservation stamp to hunt and fish in Wyoming. (See page 6) See page 18 for more information. wgfd.wyo.gov Wyoming Hunting Regulations | 1 CONTENTS Access on Lands Enrolled in the Department’s Walk-in Areas Elk or Hunter Management Areas .................................................... 4 Hunt area map ............................................................................. 46 Access Yes Program .......................................................................... 4 Hunting seasons .......................................................................... 47 Age Restrictions ................................................................................. 4 Characteristics ............................................................................. 47 Antelope Special archery seasons.............................................................. 57 Hunt area map ..............................................................................12 Disabled hunter season extension.............................................. 57 Hunting seasons ...........................................................................13 Elk Special Management Permit ................................................. 57 Characteristics ..............................................................................13 Youth elk hunters........................................................................ -

View Draft Regulation
Chapter 5, Antelope Hunting Seasons At the time of this filing, the 2020 antelope harvest information is not yet available to the Department. Individual hunt area regular hunting season dates, special archery hunting season dates, hunt area limitations, license types and license quotas may be modified after harvest data has been evaluated. Any additional proposed changes to regular hunting season dates, special archery hunting season dates, hunt area limitations, numbers of limited quota licenses, license types, hunt area boundaries or modifications to other hunting provisions shall be made available for public comment on the Department website. An updated draft of 2021antelope hunting season proposals will also be posted to the Department website during the later portion of the public comment period. Section 4, edits have been proposed to further clarify antelope hunting season provisions for persons who qualify for and are in possession of hunting season extension permits. During the 2020 hunting season, special archery season information was repositioned within this regulation and caused some confusion among hunting season extension permit holders. The edited language in this Section is meant to clarify when a hunting season extension permit is valid. Please scroll down to view the regulation or click the down arrow for the next page. Draft 1-25-2021.2 CHAPTER 5 ANTELOPE HUNTING SEASONS Section 1. Authority. This regulation is promulgated by authority of Wyoming Statutes § 23-1-302, § 23-1-703 and § 23-2-104. Section 2. Regular Hunting Seasons. Hunt areas, season dates and limitations. Special Archery Regular Hunt License Dates Season Dates Area Type Opens Closes Opens Closes Quota Limitations 1 1 Aug. -
Deer Season Subject to the Species Limitation of Their License in the Hunt Area(S) Where Their License Is Valid As Specified in Section 2 of This Chapter
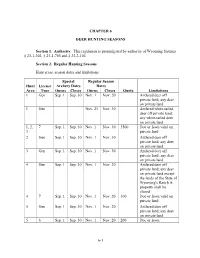
CHAPTER 6 DEER HUNTING SEASONS Section 1. Authority. This regulation is promulgated by authority of Wyoming Statutes § 23-1-302, § 23-1-703 and § 23-2-104. Section 2. Regular Hunting Seasons. Hunt areas, season dates and limitations. Special Regular Season Hunt License Archery Dates Dates Area Type Opens Closes Opens Closes Quota Limitations 1 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 20 Antlered deer off private land; any deer on private land 1 Gen Nov. 21 Nov. 30 Antlered white-tailed deer off private land; any white-tailed deer on private land 1, 2, 7 Sep. 1 Sep. 30 Nov. 1 Nov. 30 3500 Doe or fawn valid on 3 private land 2 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 30 Antlered deer off private land; any deer on private land 3 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 30 Antlered deer off private land; any deer on private land 4 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 20 Antlered deer off private land; any deer on private land except the lands of the State of Wyoming's Ranch A property shall be closed 4 7 Sep. 1 Sep. 30 Nov. 1 Nov. 20 300 Doe or fawn valid on private land 5 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 20 Antlered deer off private land; any deer on private land 5 6 Sep. 1 Sep. 30 Nov. 1 Nov. 20 200 Doe or fawn 6-1 6 Gen Sep. 1 Sep. 30 Nov. 1 Nov. 20 Antlered deer off private land; any deer on private land 7 Gen Sep. -

Carbon County DRAFT Natural Resource Management Plan
FEBRUARY 16, 2021 Carbon County DRAFT Natural Resource Management Plan Natural Resource Management Plan Y2 Consultants, LLC & Falen Law Offices (Intentionally Left Blank) Natural Resource Management Plan Y2 Consultants, LLC & Falen Law Offices CONTENTS ACRONYMS ............................................................................................................................... III LIST OF FIGURES ...................................................................................................................... VII LIST OF TABLES ......................................................................................................................... IX CHAPTER 1: INTRODUCTION .....................................................................................................10 1.1 PURPOSE ............................................................................................................................10 1.2 STATUTORY REQUIREMENTS AND LEGAL FRAMEWORK ...................................................................11 1.3 CARBON COUNTY NATURAL RESOURCE MANAGEMENT PLAN PROCESS ..............................................15 1.4 CREDIBLE DATA ....................................................................................................................19 CHAPTER 2: CUSTOM AND CULTURE ........................................................................................21 2.1 COUNTY INTRODUCTION AND OVERVIEW ....................................................................................21 2.2 CULTURAL/HERITAGE/PALEONTOLOGICAL -

ARR13-009.Pdf
STATEMENT OF REASON WYOMING GAME AND FISH COMMISSION W.S. § 23-1-302 directs and empowers the Commission to fix seasons and bag limits, open, shorten or close seasons on any species or sex of wildlife except predatory animals, predacious birds, protected animals and protected birds. W.S. § 23-2-107 empowers the Commission to promulgate reasonable rules and regulations regulating wild bison licenses and the management of wild bison. The Commission proposes to amend Wyoming Game and Fish Commission Regulations listed below to establish annual hunting seasons, limitations and bag limits. The 2012 big game harvest information is not available at the time these draft regulations are filed and made available for public comment. Any additional proposed changes to season dates, numbers and types of licenses and hunt area boundaries will be made available to the public for comment during all public meetings held around the state. Chapter 2, General Hunting Regulation Chapter 5, Antelope Hunting Seasons Chapter 6, Deer Hunting Seasons Chapter 7, Elk Hunting Seasons Chapter 8, Moose Hunting Seasons Chapter 9, Bighorn Sheep Hunting Seasons Chapter 11, Sage Grouse Hunting Seasons Chapter 12, Blue and Ruffed Grouse Hunting Seasons Chapter 13, Partridge Hunting Seasons Chapter 15, Wild Bison Recreational Hunting Season Chapter 17, Small Game Hunting Seasons Chapter 18, Pheasant Hunting Seasons Chapter 19, Sharp-Tailed Grouse Hunting Seasons Chapter 20, Wild Turkey Fall And Spring Hunting Seasons Chapter 24, Mountain Goat Hunting Seasons Chapter 39, Early Migratory Game Bird Hunting Seasons Chapter 48, Light Goose Conservation Order Chapter 2, Section 4. The Department is currently evaluating the number of doe/fawn licenses a person may apply for and receive for certain Hunt Areas. -

NORTH PLATTE FLY FISHING RANCH Alcova, Natrona County, Wyoming
Specializing in Farm, Ranch, Recreational & Auction Properties Proudly Presents NORTH PLATTE FLY FISHING RANCH Alcova, Natrona County, Wyoming Ten miles of North Platte River frontage offering world-class fly fishing Photo Courtesy of Jason Ostrander, Grey Reef Anglers & Wingshooting LOCATION & ACCESS The North Platte Fly Fishing Ranch is located near Alcova, Wyoming southwest of Casper on Wyoming Highway 220. To access the south side of the river, travel approximately 26 miles from SW Wyoming Boulevard to Gray Reef Road and turn left; proceed on Gray Reef Road for approximately 0.8 miles to the Gray Reef Dam. Vehicle access to the ten miles of river being offered for sale will be via an easement granted by the Seller to the new owner of the river frontage. Numerous public access points along the north bank of the North Platte River allow access for the new owner to all ten miles of river frontage along the south bank of the river. The following are several towns and cities that are easily accessible from the North Platte Fly Fishing Ranch. Casper, Wyoming (population 55,316) 27 miles northeast Douglas, Wyoming (population 6,120) 81 miles east Lander, Wyoming (population 7,487) 125 miles west Cheyenne, Wyoming (population 59,466) 192 miles southeast Denver, CO Metro Area (population 3,277,309) 269 miles south Rapid City, South Dakota (population 67,956) 284 miles northeast Jackson, Wyoming (population 9,577) 286 miles west North Platte Fly Fishing Ranch Page 2 www.ClarkLandBrokers.com Photo Courtesy of Jason Ostrander, Grey Reef Anglers & Wingshooting SIZE & DESCRIPTION World-class fly fishing is found throughout the ten miles of North Platte River that make up the North Platte Fly Fishing Ranch. -

ARR17-027.Pdf
STATEMENT OF REASONS WYOMING GAME AND FISH COMMISSION CHAPTER 28 REGULATION GOVERNING BIG OR TROPHY GAME ANIMAL OR GAME BIRD DAMAGE CLAIMS W.S. § 23-1-302 (a) (xxii) provides the Commission the authority to promulgate such orders as the Commission considers necessary to carry out the intent of this act. W.S. § 23-1-901 stipulates the process for owners of damaged property to report damages caused by big or trophy game or game birds. House Enrolled Act #117, Sixty-Fourth Legislature of the State of Wyoming, 2017 General Session, provides for amendments to W.S. § 23-1-901 (g) and (h) which empowers the Commission to establish rules to allow a property owner to request that an area of land adjacent to the area described in W.S. § 23-1-101 (a) (xii) (B) (I) or (II) be designated as an area where property owners are eligible for damage compensation for damage caused by gray wolves. This act is effective July 1, 2017. Throughout this draft regulation, language has been added to various Sections to include “gray wolf in accordance with W. S. § 23-1-901 (g) and (h)”. This reference allows a claimant to request damage compensation for gray wolf damage to livestock on land adjacent to the area described in W.S. § 23-1-101 (a) (xii) (B) (I) or (II) in accordance with statutory and regulatory provisions. “Gray Wolf” will also be added to the title of this regulation. Section 2, Definitions. The definitions of “Claimant, Confirmed by the Department or its Representative, Consequential Damages, Damage, Investigated by the Department and Verified Claim” are being modified to include new statutory provisions. -

SEVEN X RANCH Casper, Natrona County, Wyoming
Specializing in Farm, Ranch, Recreational & Auction Properties Proudly Presents SEVEN X RANCH Casper, Natrona County, Wyoming A well-balanced cow-calf grazing operation with an irrigated feed base, year-round live water, and excellent, well-designed, improvements. LOCATION & ACCESS The Seven X Ranch is located approximately 20 miles southwest of Casper, Wyoming at 14355 Clark Road. There is year-round access from paved Wyoming Highway 220. To access the ranch from Casper, drive southwest from Casper on WY-220 for approximately 19 miles to Clark Road, turn left onto Clark Road for one mile which leads to the main entrance of the Seven X Ranch. Several towns and cities in proximity to the property include: • Laramie, Wyoming (population 30,816) 129 miles south • Cheyenne, Wyoming (population 59, 466) 199 miles southeast • Casper, Wyoming (population 59,628) 20 miles northeast • Glenrock, Wyoming (population 2,591) 45 miles northeast • Rawlins, Wyoming (population 8,858) 97 miles south • Fort Collins, Colorado (population 143,986) 193 miles southeast • Denver, Colorado (population 701,621) 258 miles southeast Seven X Ranch Page 2 www.ClarkLandBrokers.com DESCRIPTION 2,151± Acres Deeded 780± Acres BLM Lease 720± State of Wyoming Lease 3,651± Total Contiguous Acres The ranch consists of pivot-irrigated acres, flood-irrigated pastures, dryland grazing, and well- developed improvements for livestock, equipment, and occupants. The property is fenced with four and five strands of barbed wire. It is cross fenced into nine pastures for rotational grazing. The terrain of the ranch consists of irrigated meadows, creek bottom riparian zone with gentle sloping grass hills, and rolling hills of native grass pasture, approximately 300 acres of which are flood-irrigated in the spring. -

ARR14-010.Pdf
STATEMENT OF REASONS WYOMING GAME AND FISH COMMISSION W.S. §23-1-302 directs and empowers the Commission to fix seasons and bag limits, open, shorten or close seasons on any species or sex of wildlife except predatory animals, predacious birds, protected animals and protected birds. W.S. §23-2-107 empowers the Commission to promulgate reasonable rules and regulations regulating wild bison licenses and the management of wild bison. The Commission proposes to amend Wyoming Game and Fish Commission Regulations listed below to establish annual hunting seasons, limitations and bag limits for the 2014 hunting seasons. The 2013 big game harvest information is not available at the time these draft regulations are filed and made available for public comment. Any additional proposed changes to season dates, numbers of limited quota licenses, types of licenses, hunt area boundaries or other hunting provisions will be made available for public comment during all public meetings held around the state. Chapter 2, General Hunting Regulation Chapter 5, Antelope Hunting Seasons Chapter 6, Deer Hunting Seasons Chapter 7, Elk Hunting Seasons Chapter 8, Moose Hunting Seasons Chapter 9, Bighorn Sheep Hunting Seasons Chapter 11, Sage Grouse Hunting Seasons Chapter 12, Blue and Ruffed Grouse Hunting Seasons Chapter 13, Partridge Hunting Seasons Chapter 15, Wild Bison Recreational Hunting Season Chapter 17, Small Game Hunting Seasons Chapter 18, Pheasant Hunting Seasons Chapter 19, Sharp-Tailed Grouse Hunting Seasons Chapter 20, Wild Turkey Fall And Spring Hunting Seasons Chapter 24, Mountain Goat Hunting Seasons Chapter 39, Early Migratory Game Bird Hunting Seasons Chapter 48, Light Goose Conservation Order Chapter 2, General Hunting Regulation, Section 4. -
Carbon County Natural Resource Management Plan
JULY 6, 2021 Carbon County Natural Resource Management Plan Natural Resource Management Plan Y2 Consultants, LLC & Falen Law Offices (Intentionally Left Blank) Natural Resource Management Plan Y2 Consultants, LLC & Falen Law Offices CONTENTS ACRONYMS .......................................................................................................................... III LIST OF FIGURES .................................................................................................................. VII LIST OF TABLES ..................................................................................................................... IX CHAPTER 1: INTRODUCTION ................................................................................................ 10 1.1 PURPOSE................................................................................................................................. 10 1.2 STATUTORY REQUIREMENTS AND LEGAL FRAMEWORK ..................................................................... 11 1.3 CARBON COUNTY NATURAL RESOURCE MANAGEMENT PLAN PROCESS ............................................... 15 1.4 CREDIBLE DATA ........................................................................................................................ 19 CHAPTER 2: CARBON COUNTY CUSTOM AND CULTURE ........................................................ 21 2.1 COUNTY INTRODUCTION AND OVERVIEW ...................................................................................... 21 2.2 CULTURAL/HERITAGE/PALEONTOLOGICAL RESOURCES -

Clark & Associates Land Brokers
Specializing in Farm, Ranch, Recreational & Auction Properties Proudly Presents OPEN AT FARM Casper, Natrona County, Wyoming The Open AT Farm is a turn-key hay farm, cattle operation and gravel pit all bordered by the North Platte River. LOCATION & ACCESS The Open AT Farm is located approximately 20 miles southwest of Casper, Wyoming. There is year-round access from paved Wyoming Highway 220 leading to the private driveway. To access the farm from Casper, travel west on Highway 220 for 19 miles. The destination is on your right at 16550 State Highway 220. Several towns and cities in proximity to the property include: Laramie, Wyoming (population 30,816) 129 miles south Cheyenne, Wyoming (population 59, 466) 199 miles southeast Casper, Wyoming (population 59,628) 20 miles northeast Glenrock, Wyoming (population 2,591) 45 miles northeast Rawlins, Wyoming (population 8,858) 97 miles south Fort Collins, Colorado (population 143,986) 193 miles southeast Denver, Colorado (population 701,621) 258 miles southeast Open AT Farm Page 2 www.ClarkLandBrokers.com SIZE & DESCRIPTION 1,011.74+ Acres Deeded 393.95+ State of Wyoming Lease 79.93+ Acres BLM Lease 1,515.71+ Total Contiguous Acres The farm consists of four Zimmatic pivots, flood irrigation, and a gravel pit. The property is fenced with four and five strands of barbed wire. It is cross fenced into six pastures for rotational grazing. The terrain of the farm consists of irrigated meadows, creek bottom, and over a mile of North Platte River frontage with gentle sloping grass hills. There are mature box-elder, elm, cottonwood and willow trees which provide excellent habitat for the wildlife and winter protection for the livestock. -

Summary Appraisal Report the Properties Located at Alcova/Pathfinder Reservoir Alcova, Wyoming 82620 Book Two - Analysis
SUMMARY APPRAISAL REPORT THE PROPERTIES LOCATED AT ALCOVA/PATHFINDER RESERVOIR ALCOVA, WYOMING 82620 BOOK TWO - ANALYSIS Prepared For Mr. Mike Haigler, Road, Bridge and Parks Director Board of County Commissioners Natrona County Date of Appraisal - January 30, 2012 Effective Date - December 1, 2011 Appraised By Edward J. Holthouse, CRS, CCIM Certified General Real Estate Appraiser SUBSECTION “A” APPRAISAL ANALYSIS ON THE ALCOVA CABIN SITES 1-100 ALCOVA RESERVOIR NATRONA COUNTY, WYOMING The following analysis is only part of the “Total Appraisal Report”, referred herein as Appraisal Report No. 6705, which includes both Book One – Basic Data Section plus addenda and Book Two – Parcel Analysis. Except as hereinafter provided, the Client may distribute copies of the “total appraisal report” in its entirety to such third parties as they may select; however, selected portions of this appraisal report shall not be given to third parties without the prior written consent of the signatories of this appraisal report. Holthouse Appraisal Group Analysis – 6705 Page A - 1 Holthouse Appraisal Group The Appraisal Firm of Phone Number - 307-265-7908 Edward J. Holthouse and Associates Fax Number - 307-265-3754 139 West Second Street, Suite 3E E-Mail - [email protected] PO Box 1747 Casper, Wyoming 82602-1747 January 30, 2012 Mr. Mike Haigler, Director Natrona County Road, Bridge and Parks Department 538 SW Wyoming Boulevard Mills, WY 82644 Dear Mr. Haigler: In fulfillment of our agreement, I have performed an appraisal on the real property, land only, known as Alcova Cabin Sites 1-100, Alcova Reservoir, Natrona County, Wyoming, herein referred to as the "subject property".