Redalyc.Palaeontology of the Upper Turonian Paralic Deposits of The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Groupement De Gendarmerie Departementale De La Dordogne
GROUPEMENT DE GENDARMERIE DEPARTEMENTALE DE LA DORDOGNE Conseiller Réserves Commandant de Groupement Commandant en Second COL NICOT Lionel COL (ER) THIRIOT Philippe LCL RAYNE Serge Section Commandement Centre Soutien Automobile Gendarmerie ADC CHARLES Sandrine MAJ COUSTY Jean-Michel Chauffeur commandant de groupement Officier Adjoint Commandement GND GALLI Stéphane LCL CAUREZ José Officier Adjoint Groupe Soutien Service Logistique Finances Ressources Humaines ADJ LEFEVRE Jean-François CNE DIGOT Sébastien Brigade Départementale de Renseignements et d'Investigations Judiciaires Service Affaires Immobilières MAJ DUBREUIL Jean-Louis Officier Adjoint Police Judiciaire MAJ GIRARDEAU Christophe LCL SANCHEZ Michel Cellule Identification Criminelle Numérique Service Administration du Personnel M. BOUVIER Didier MAJ SICARD Jean-François Officier Adjoint Renseignement – Recrutement Section des Systèmes d'Information et de Cellule Réserve Départementale CNE LE DREN Marc Communication Cellule Renseignement – Recrutement MME TRONCHE Chantal MAJ DOCHE Thierry MDC GARLOPEAU Géraldine Officier Adjoint Centre Opérationnel – Officier Section Administration du Personnel Adjoint Prévention CNE AMOROSO Joseph Cellule Prévention Technique Malveillance MJR CORBASSON Daniel Chef de Quart Opérateur ADJE MORAND Sylvie COMPAGNIE COMPAGNIE COMPAGNIE COMPAGNIE ESCADRON DEPARTEMENTAL BERGERAC NONTRON PERIGUEUX SARLAT SECURITE ROUTIERE CNE COURTON Damien CNE MERCIER Emilie CEN ANDRIEUX Sébastien CEN TETU Matthieu CNE DERUDDER Jean-Claude CNE LAURENT Patrice CNE VINCENT -

ZRR : Exonération De Taxe Foncière Sur Les Propriétés Bâties (TFPB)
ZRR : exonération de Taxe Foncière sur les Propriétés Bâties (TFPB) DDFIP Présentation du dispositif Les entreprises qui créent ou reprennent une entreprise en difficulté, dans une ZRR (Zone de revitalisation rurale), peuvent bénéficier d'une exonération de taxe foncière sur les propriétés bâties. Sont concernées les entreprises de moins de 11 salariés : exerçant une activité industrielle, commerciale, artisanale ou libérale, créées ou reprises en ZRR jusqu'au 31/12/2022. Conditions d'attribution A qui s’adresse le dispositif ? Entreprises éligibles$$Critères d’éligibilité Pour quel projet ? Présentation des projets Dépenses concernées Quelles sont les particularités ? Entreprises inéligibles$$Critères d’inéligibilité$$Dépenses inéligibles Montant de l'aide L'exonération est comprise entre 2 et 5 ans. L'exonération porte sur la totalité de la part revenant à chaque collectivité territoriale, groupement doté d'une fiscalité propre ou établissement public ayant pris une délibération. Seuls les services fiscaux sont compétents pour juger de l'éligibilité de l'entreprise au présent dispositif. Informations pratiques L'entreprise doit déclarer, dans les 15 jours de la signature de l'acte, son acquisition au service des impôts. DDFIP ZRR : exonération de Taxe Foncière sur les Propriétés Bâties (TFPB) Page 1 sur 5 Le délai de 5 ans est calculé à partir du 01/01 de la première année au titre de laquelle l'exonération de taxe foncière a été accordée. Critères complémentaires Création avant le 31 décembre 2020. Effectif de moins de 11 salariés. -

BULLETIN MUNICIPAL N° 16 Le Mot Du Maire
BULLETIN MUNICIPAL N° 16 Le mot du Maire Chers concitoyens, Je souhaiterais tout d’abord remercier toutes celles et tous ceux d’entre vous qui, lors des élections municipales du 23 mars dernier nous ont accordé leur confiance pour conduire à nouveau pendant 6 ans, l’avenir de notre commune. Permettez-moi de voir dans le résultat des urnes, le signe d’une certaine reconnaissance du travail accompli depuis 2008. A mes collègues qui ont cessé leur mandat, j’exprime mes remerciements pour leur dévouement pendant les années passées au service de notre collectivité. Forte de cette confiance, c’est une équipe légèrement renouvelée (trois nouveaux élus sont entrés au conseil municipal) qui sans tarder s’est emparée des projets en cours afin que nous restions une commune moderne, attractive et tournée vers l’avenir. Les années qui s’ouvrent ne seront pas de tout repos, nous subissons en effet une crise économique importante depuis plusieurs années. La réforme territoriale, la mise en place d’un nouveau plan local d’urbanisme intercommunal et la réduction des dotations de l’Etat nous obligent d’ores et déjà à penser l’action publique autrement. Il faudra faire aussi bien mais avec moins de moyens. Nous devrons donc redoubler de créativité et d’énergie pour défendre notre village. Je reste néanmoins optimiste car j’ai l’intime conviction que la ruralité a de nombreux atouts et donc de l’avenir. Je sais aussi que nous pourrons toujours compter, certes dans la mesure du possible, sur le soutien financier de l’Etat, du Département et de la Région. -

Servitudes D'utilite Publique De La Communaute De Communes Du Pays De Fenelon
SERVITUDES D'UTILITE PUBLIQUE DE LA COMMUNAUTE DE COMMUNES DU PAYS DE FENELON NOM COMMUNE Code Catégorie DESIGNATION NOM TEXTE DATE TEXTE CONSERVATION DU PATRIMOINE ARCHIGNAC AC 1 - Eglise Arrêté du 29/11/48 CULTUREL : Monuments historiques UTILISATION DE CERTAINES Arrêté RESSOURCES ET EQUIPEMENTS : liaison hertzienne Sarlat/Croix ARCHIGNAC PT 2 Préfectoral 02/02/78 Télécommunications-protection contre les d'Allon du obstacles UTILISATION DE CERTAINES ARCHIGNAC I 4 RESSOURCES ET EQUIPEMENTS : Diverses lignes MT et BT Convention Energie électrique UTILISATION DE CERTAINES BORREZE I 4 RESSOURCES ET EQUIPEMENTS : Diverses lignes MT + BT Convention Energie électrique Arrêté CONSERVATION DU PATRIMOINE BORREZE AS 1 Puits de Borrèze ou Canteranne Préfectoral 01/02/80 NATUREL : Conservation des eaux du Arrêté CONSERVATION DU PATRIMOINE BORREZE AS 1 Forage de Peyrenègre Préfectoral 26/02/96 NATUREL : Conservation des eaux du CONSERVATION DU PATRIMOINE CALVIAC EN PERIGORD AC 1 - Eglise Arrêté du 18/09/70 CULTUREL : Monuments historiques CONSERVATION DU PATRIMOINE - Eglise d Aillac (commune de CALVIAC EN PERIGORD AC 1 Arrêté du 21/10/70 CULTUREL : Monuments historiques CARSAC AILLAC) CONSERVATION DU PATRIMOINE - Site de la Dordogne de Vitrac à CALVIAC EN PERIGORD AC 2 Arrêté du 14/10/69 CULTUREL : Monuments naturels et sites Cazoulès PLAN D EXPOSITION AUX RISQUES Arrêté NATURELS PREVISIBLES : PLAN D CALVIAC EN PERIGORD PM 1 PPRinondation de la Dordogne Préfectoral 15/04/11 EXPOSITION AUX RISQUES NATURELS du PREVISIBLES - Cheminée du 14ème CONSERVATION -

Bulletin Municipal
Année 2017 Edition janvier 2018 Bulletin municipal 2017 Consultable sur : www.carlux24.fr Page 2 Année 2017 Bulletin municipal CARLUX joli village du Périgord noir où restent des traces des guerres de Cent ans et de religion, se situe dans la vallée de la DORDOGNE. Altitude : 85 à 260 mètres 647 habitants 1,500 km de la rivière Dordogne. 15 km de Sarlat (Pays de La Boétie) 8 km du château de Fénelon 15 km de l’autoroute Paris Toulouse (Autoroute A20) 30 km de l’axe routier Bordeaux-Clermont-Ferrand-Lyon (Autoroute A89) 15 km de la gare S.N.C.F. de Souillac (ligne Paris-Toulouse) 20 km de l’aéroport Brive-vallée-Dordogne (http://www.aeroport-brive-vallee-dordogne.com/) Piste cyclable voie verte vélo route Sarlat-Cazoulès sur la commune. Équipements collectifs Foyer Seyral salle de réunion Salle polyvalente équipée (salle des fêtes) Place des Platanes (Boules, Spectacles plein air, …) Terrain de sport Chemins pédestres : GR6, nombreux circuits de randonnées (PDIPR) Parcours VTT balisés Zone Artisanale de Rouffillac Commerces Commerces itinérants Libre service Proxi Pizza Big Good Halle paysanne Poissonnerie Boulangerie Boucherie Cafés restaurants Hôtels Services publics Agence Postale communale Maison du tourisme Pays de Fénelon Gare de Carlux Espace DOISNEAU Maison Relais des services publics Médecin Ostéopathes Psychanalyste Gendarmerie Année 2017 Page 3 LE MOT DU MAIRE 4 LA VIE MUNICIPALE 5 Le Conseil municipal 2014-2020 5-6 Les services municipaux 6 Le budget 2017 L’état civil 7 Le mot des gendarmes 8 Listes électorales, cartes d’identité, -

Commission Implementing Decision (Eu) 2015
3.12.2015 EN Official Journal of the European Union L 317/37 COMMISSION IMPLEMENTING DECISION (EU) 2015/2239 of 2 December 2015 concerning certain protective measures in relation to highly pathogenic avian influenza of subtypes H5N1 and H5N2 in France (notified under document C(2015) 8755) (Only the French text is authentic) (Text with EEA relevance) THE EUROPEAN COMMISSION, Having regard to the Treaty on the Functioning of the European Union, Having regard to Council Directive 89/662/EEC of 11 December 1989 concerning veterinary checks in intra- Community trade with a view to the completion of the internal market (1), and in particular Article 9(4) thereof, Having regard to Council Directive 90/425/EEC of 26 June 1990 concerning veterinary and zootechnical checks applicable in intra-Community trade in certain live animals and products with a view to the completion of the internal market (2), and in particular Article 10(4) thereof, Whereas: (1) Avian influenza is an infectious viral disease in birds, including poultry. Infections with avian influenza viruses in domestic poultry cause two main forms of that disease that are distinguished by their virulence. The low pathogenic form generally only causes mild symptoms, while the highly pathogenic form results in very high mortality rates in most poultry species. That disease may have a severe impact on the profitability of poultry farming. (2) Avian influenza is mainly found in birds, but under certain circumstances infections can also occur in humans even though the risk is generally very low. (3) In the event of an outbreak of avian influenza, there is a risk that the disease agent might spread to other holdings where poultry or other captive birds are kept. -

C99 Official Journal
Official Journal C 99 of the European Union Volume 63 English edition Information and Notices 26 March 2020 Contents II Information INFORMATION FROM EUROPEAN UNION INSTITUTIONS, BODIES, OFFICES AND AGENCIES European Commission 2020/C 99/01 Non-opposition to a notified concentration (Case M.9701 — Infravia/Iliad/Iliad 73) (1) . 1 2020/C 99/02 Non-opposition to a notified concentration (Case M.9646 — Macquarie/Aberdeen/Pentacom/JV) (1) . 2 2020/C 99/03 Non-opposition to a notified concentration (Case M.9680 — La Voix du Nord/SIM/Mediacontact/Roof Media) (1) . 3 2020/C 99/04 Non-opposition to a notified concentration (Case M.9749 — Glencore Energy UK/Ørsted LNG Business) (1) . 4 IV Notices NOTICES FROM EUROPEAN UNION INSTITUTIONS, BODIES, OFFICES AND AGENCIES European Commission 2020/C 99/05 Euro exchange rates — 25 March 2020 . 5 2020/C 99/06 Summary of European Commission Decisions on authorisations for the placing on the market for the use and/or for use of substances listed in Annex XIV to Regulation (EC) No 1907/2006 of the European Parliament and of the Council concerning the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH) (Published pursuant to Article 64(9) of Regulation (EC) No 1907/2006) (1) . 6 EN (1) Text with EEA relevance. V Announcements PROCEDURES RELATING TO THE IMPLEMENTATION OF COMPETITION POLICY European Commission 2020/C 99/07 Prior notification of a concentration (Case M.9769 — VW Group/Munich RE Group/JV) Candidate case for simplified procedure (1) . 7 OTHER ACTS European Commission 2020/C 99/08 Publication of a communication of approval of a standard amendment to a product specification for a name in the wine sector referred to in Article 17(2) and (3) of Commission Delegated Regulation (EU) 2019/33 . -

Guide Des Itinéraires
1 2 3 4 VTT Le Bugue - Les Eyzies Le Bugue - Saint-Cirq Le Bugue - Audrix - Campagne Le Bugue - Campagne itinéraires des Le Bugue ou les Eyzies de Tayac-Sireuil, Le Bugue, aire de Camping-car Le Bugue, aire de Camping-car Le Bugue, aire de Camping-car Départ : aire de Camping-car Départ : Départ : Départ : Guide 22,3 km + 450 m 28,6 km + 580 m 10,5 km + 130 m 38,3 km + 950 m Distance : - Dénivelé : Distance : - Dénivelé : Distance : - Dénivelé : Distance : - Dénivelé : Le bourg d’Audrix, le gouffre de Proumeyssac, Le bourg d’Audrix, le gouffre de Proumeyssac, Saint-Cirq, la grotte du Sorcier, la Ferrassie et son La rivière Vézère, le Bugue. à voir : à voir : à voir : à voir : le hameau de Péchalifour, Lussac, les Eyzies de Tayac et tous le hameau de Péchalifour, la forêt de Campagne et son point gisement préhistorique, le Bugue, la grotte de Bar-Bahaut. ces sites préhistoriques. de vue, les cluzeaux et le château de Campagne, le Bugue. 5 6 7 8 Rouffignac - le Moustier Le tour de Rouffignac Rouffignac - l ’Herm Rouffignac - Mauzens et Miremont Rouffignac St Cernin de Reilhac, Salle Rouffignac St Cernin de Reilhac, Salle Rouffignac St Cernin de Reilhac, Salle Rouffignac St Cernin de Reilhac, Salle Départ : Départ : Départ : OmnisportsDépart : (ESES) du Complexe Sportif Omnisports (ESES) du Complexe Sportif Omnisports (ESES) du Complexe Sportif Omnisports (ESES) du Complexe Sportif 42,6 km + 1 395 m 5 km + 60 m 29,3 km + 661 m 26,6 km + 565 m Distance : Dénivelé : Distance : - Dénivelé : Distance : - Dénivelé : Distance : - Dénivelé : - Une très belle vue sur la vallée de la Vézère, ses Les alentours de Rouffignac avec les 5 routes Traversée de Plazac, point de vue sur Rouffignac Passage à proximité du village de St-Cernin, joli à voir : à voir : à voir : à voir : méandres et sur la Maison Forte de Reignac. -

Présentation A4
Annexe n°2 à l'arrêté préfectoral n° 110548 du 25 mai 2011 Le risque sismique en Dordogne Zone 2 - niveau faible BUSSEROLLES CHAMPNIERS Zone 1 - niveau trés faible ET-REILHAC BUSSIERE BADIL SAINT-BARTHELEMY DE-BUSSIERE PIEGUT PLUVIERS SOUDAT VARAIGNES SAINT ETOUARS ESTEPHE TEYJAT FIRBEIX ABJAT SAINT-PIERRE AUGIGNAC SUR-BANDIAT DE-FRUGIE LE BOURDEIX JAVERLHAC-ET LA-CHAPELLE-ST-ROBERT MIALET SAINT-MARTIN LE-PIN SAVIGNAC DE-NONTRON LA COQUILLE CHAMPS SAINT-SAUD SAINT-PRIEST HAUTEFAYE ROMAIN LACOUSSIERE LES-FOUGERES LUSSAS-ET NONTRONNEAU NONTRON BEAUSSAC CONNEZAC ST-MARTIAL DE-VALETTE JUMILHAC-LE-GRAND LES GRAULGES ST-PARDOUX CHALEIX LA-RIVIERE SCEAU RUDEAU ST-ANGEL LADOSSE PUYRENIER ST-FRONT SAINT-PAUL SUR-NIZONNE LA-ROCHE SAINTE-CROIX SAINT-JORY DE-MAREUIL ST-SULPICE CHAMPEAUX-ET-LA MILHAC-DE DE-CHALAIS SAINT-FRONT DE-MAREUIL CHAPELLE-POMMIER NONTRON LA ROCHEBEAUCOURT LA-RIVIERE ET-ARGENTINE LA CHAPELLE SARLANDE MONTMOREAU ST-MARTIN-DE VIEUX FRESSENGEAS THIVIERS MAREUIL MAREUIL NANTHEUIL CHAMPAGNE QUINSAC SARRAZAC ET-FONTAINE MONSEC SAINT-CREPIN SAINT DE-RICHEMONT PANCRACE ST-ROMAIN-ET ANGOISSE GOUT VILLARS VENDOIRE ST-CLEMENT PAYZAC ROSSIGNOL ST-FELIX-DE ST-JEAN BOURDEILLES DE-COLE LA CHAPELLE CHAMPAGNAC MONTABOURLET CANTILLAC DE-BELAIR LA CHAPELLE NANTHIAT ST-SULPICE GRESIGNAC CHERVAL LEGUILLAC-DE D'EXCIDEUIL DUSSAC CERCLES ST-PIERRE LA GONTERIE EYZERAC NANTEUIL-AURIAC DE-COLE BOULOUNEIX SAVIGNAC DE-BOURZAC LA CHAPELLE CORGNAC LANOUAILLE LEDRIER FAUCHER SUR-L'ISLE SAINT-MARTIAL LA TOUR CERCLES SAINT-CYR VAUNAC VIVEYROL BLANCHE ST-JULIEN-DE -

Liste Des Villes Et Villages Du Département De La Dordogne
Liste des villes et villages du département de la Dordogne Communes du departement de la Dordogne en A Abjat-sur-Bandiat Agonac Ajat Allas-les-Mines Allemans Alles-sur-Dordogne Angoisse Anlhiac Annesse-et-Beaulieu Antonne-et-Trigonant Archignac Atur Aubas Audrix Augignac Auriac-du-Périgord Azerat Communes du departement de la Dordogne en B Badefols-d'Ans Badefols-sur-Dordogne Baneuil Bardou Bars Bassillac Bayac Beaumont-du-Périgord Beaupouyet Beauregard-de-Terrasson Beauregard-et-Bassac Beauronne Beaussac Beleymas Belvès Berbiguières Bergerac Bertric-Burée Besse Beynac-et-Cazenac Bézenac Biras Biron Blis-et-Born Boisse Boisseuilh Bonneville-et-Saint-Avit-de-Fumadières Borrèze Bosset Bouillac Boulazac Bouniagues Bourdeilles Bourg-des-Maisons Bourg-du-Bost Bourgnac Bourniquel Bourrou Bouteilles-Saint-Sébastien Bouzic Brantôme Breuilh Brouchaud Bussac Busserolles Bussière-Badil Communes du departement de la Dordogne en C Calès Calviac-en-Périgord Campagnac-lès-Quercy Campagne Campsegret Cantillac Capdrot Carlux Carsac-Aillac Carsac-de-Gurson Carves Castelnaud-la-Chapelle Castels Cause-de-Clérans Cazoulès Celles Cénac-et-Saint-Julien Cendrieux Cercles Chalagnac Chaleix Champagnac-de-Belair Champagne-et-Fontaine Champcevinel Champeaux-et-la-Chapelle-Pommier Champniers-et-Reilhac Champs-Romain Chancelade Chantérac Chapdeuil Chassaignes Château-l'Évêque Châtres Chavagnac Chenaud Cherval Cherveix-Cubas Chourgnac Cladech Clermont-d'Excideuil Clermont-de-Beauregard Colombier Coly Comberanche-et-Épeluche Condat-sur-Trincou Condat-sur-Vézère Conne-de-Labarde -
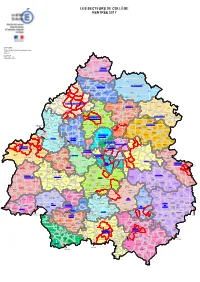
Mise En Page
LES SECTEURS DE COLLÈGE RENTRÉE 2017 D6/Informatique Source : Ramsese Fichier : ./SecteurCLG/2017-2018/SecteurCLG.wor 17/05/2017 Mise à jour : BD CARTO® © IGN - Paris - 2004 Reproduction interdite CHAMPNIERS BUSSEROLLES ET-REILHAC BUSSIERE SAINT BADIL BARTHELEMY PIEGUTDE-BUSSIERE SOUDAT VARAIGNES PLUVIERS ST-ESTEPHE ETOUARS TEYJAT FIRBEIX AUGIGNAC ST-PIERRE DE-FRUGIE LE BOURDEIX ABJAT-SUR BANDIAT JAVERLHAC ST-MARTIN MIALET ET-LA-CHAPELLE LE-PIN ST-ROBERT SAVIGNAC DE NONTRON CHAMPS HAUTEFAYE ST-PRIEST ROMAIN ST-SAUD LA COQUILLELES-FOUGERES LUSSAS-ET NONTRON LACOUSSIERE NONTRONNEAU CONNEZAC ST-MARTIAL BEAUSSAC DE-VALETTE JUMILHAC LES GRAULGES LE GRAND ST PARDOUX RUDEAU LA RIVIERE CHALEIX LADOSSE SCEAU PUYRENIER ST-ANGEL ST-PAUL STE-CROIX ST-FRONT LA ROCHE- LA-ROCHE DE SAINT-SULPICE SUR-NIZONNE ST-JORY BEAUCOURT MILHAC-DE MAREUIL DE-MAREUIL DE-CHALAIS ET-ARGENTINE NONTRON CHAMPEAUX-ET ST-FRONT LA-CHAPELLE-POMMIER LA-RIVIERE LA CHAPELLE MAREUIL VIEUX-MAREUIL ST-MARTIN-DE MONTMOREAU FRESSENGEAS SARLANDE CHAMPAGNE MAREUIL EN PÉRIGORD ET-FONTAINE ST-CREPIN DE QUINSAC SARRAZAC ST GOUT MONSEC RICHEMONT VENDOIRE ROSSIGNOL PANCRACE VILLARS THIVIERS ANGOISSE ST-FELIX DE ST-JEAN ST-ROMAIN PAYZAC BOURDEILLES CHAMPAGNAC ET NANTHEUIL LA CHAPELLE DE DE-COLE LA CHAPELLE CANTILLAC ST-CLEMENT DUSSAC NANTEUIL MONTABOURLET BELAIR GRESIGNAC AURIAC LEGUILLAC NANTHIAT CHERVAL LA GONTERIE LA DE-BOURZAC DE-CERCLES ST-PIERRE EYZERAC LA TOUR-BLANCHE BOULOUNEIX CHAPELLE DE-COLE ST-SULPICE CONDAT SAVIGNAC ST-MARTIAL FAUCHER D'EXCIDEUIL LA TOUR BLANCHE-CERCLES SUR VAUNAC -

2019 Edition Janvier 2020 Bulletin Municipal
Année 2019 Edition janvier 2020 Bulletin municipal 2019 Consultable sur : www.carlux24.fr Page 2 Année 2019 Bulletin municipal CARLUX joli village du Périgord noir où restent des traces des guerres de Cent ans et de religion, se situe dans la vallée de la DORDOGNE. Altitude : 85 à 260 mètres 639 habitants 1,500 km de la rivière Dordogne. 15 km de Sarlat (Pays de La Boétie) 8 km du château de Fénelon 15 km de l’autoroute Paris Toulouse (Autoroute A20) 30 km de l’axe routier Bordeaux-Clermont-Ferrand-Lyon (Autoroute A89) 15 km de la gare S.N.C.F. de Souillac (ligne Paris-Toulouse) 15 km de la gare S.N.C.F. de Sarlat (ligne Sarlat-Bordeaux) 20 km de l’aéroport Brive-vallée-Dordogne (http://www.aeroport-brive-vallée-dordogne.com/) Piste cyclable voie verte vélo route Sarlat-Cazoulès sur la commune. Équipements collectifs Foyer Seyral salle de réunion Salle polyvalente équipée (salle des fêtes) Place des Platanes (Boules, Spectacles plein air, …) Terrain de sport Chemins pédestres : GR6, nombreux circuits de randonnées (PDIPR) Parcours VTT balisés Commerces itinérants Commerces Pizza Big-Good Libre service Proxi Poissonnerie Halle paysanne Boucherie Boulangerie Cafés restaurants Zone Artisanale de Rouffillac Hôtels Services au public La ferme de la Chapelle Agence Postale communale Maison du tourisme Pays de Fénelon Gare de Carlux Espace DOISNEAU Maison de services au public Médecin Ostéopathes Psychanalyste Gendarmerie Année 2019 Page 3 LE MOT DU MAIRE 4 LA VIE MUNICIPALE 5 Le Conseil municipal 2014-2020 5-6 Les services municipaux 6 Le budget