Behavioral Ecology of Territorial Aggression in Uca Pugilator and Uca Pugnax
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Learning in Stomatopod Crustaceans
International Journal of Comparative Psychology, 2006, 19 , 297-317. Copyright 2006 by the International Society for Comparative Psychology Learning in Stomatopod Crustaceans Thomas W. Cronin University of Maryland Baltimore County, U.S.A. Roy L. Caldwell University of California, Berkeley, U.S.A. Justin Marshall University of Queensland, Australia The stomatopod crustaceans, or mantis shrimps, are marine predators that stalk or ambush prey and that have complex intraspecific communication behavior. Their active lifestyles, means of predation, and intricate displays all require unusual flexibility in interacting with the world around them, imply- ing a well-developed ability to learn. Stomatopods have highly evolved sensory systems, including some of the most specialized visual systems known for any animal group. Some species have been demonstrated to learn how to recognize and use novel, artificial burrows, while others are known to learn how to identify novel prey species and handle them for effective predation. Stomatopods learn the identities of individual competitors and mates, using both chemical and visual cues. Furthermore, stomatopods can be trained for psychophysical examination of their sensory abilities, including dem- onstration of color and polarization vision. These flexible and intelligent invertebrates continue to be attractive subjects for basic research on learning in animals with relatively simple nervous systems. Among the most captivating of all arthropods are the stomatopod crusta- ceans, or mantis shrimps. These marine creatures, unfamiliar to most biologists, are abundant members of shallow marine ecosystems, where they are often the dominant invertebrate predators. Their common name refers to their method of capturing prey using a folded, anterior raptorial appendage that looks superficially like the foreleg of a praying mantis. -

Cannibalism and Habitat Selection of Cultured Chinese Mitten Crab: Effects of Submerged Aquatic Vegetation with Different Nutritional and Refuge Values
water Article Cannibalism and Habitat Selection of Cultured Chinese Mitten Crab: Effects of Submerged Aquatic Vegetation with Different Nutritional and Refuge Values Qingfei Zeng 1,*, Erik Jeppesen 2,3, Xiaohong Gu 1,*, Zhigang Mao 1 and Huihui Chen 1 1 State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China; [email protected] (Z.M.); [email protected] (H.C.) 2 Department of Bioscience, Aarhus University, Vejlsøvej 25, 8600 Silkeborg, Denmark; [email protected] 3 Sino-Danish Centre for Education and Research, University of CAS, Beijing 100190, China * Correspondence: [email protected] (Q.Z.); [email protected] (X.G.) Received: 26 September 2018; Accepted: 26 October 2018; Published: 29 October 2018 Abstract: We examined the food preference of Chinese mitten crabs, Eriocheir sinensis (H. Milne Edwards, 1853), under food shortage, habitat choice in the presence of predators, and cannibalistic behavior by comparing their response to the popular culture plant Elodea nuttallii and the structurally more complex Myriophyllum verticillatum L. in a series of mesocosm experiments. Mitten crabs were found to consume and thus reduce the biomass of Elodea, whereas no negative impact on Myriophyllum biomass was recorded. In the absence of adult crabs, juveniles preferred to settle in Elodea habitats (appearance frequency among the plants: 64.2 ± 5.9%) but selected for Myriophyllum instead when adult crabs were present (appearance frequency among the plants: 59.5 ± 4.9%). The mortality rate of mitten crabs in the absence of plant shelter was higher under food shortage, primarily due to cannibalism. -

Visual Adaptations in Crustaceans: Chromatic, Developmental, and Temporal Aspects
FAU Institutional Repository http://purl.fcla.edu/fau/fauir This paper was submitted by the faculty of FAU’s Harbor Branch Oceanographic Institute. Notice: ©2003 Springer‐Verlag. This manuscript is an author version with the final publication available at http://www.springerlink.com and may be cited as: Marshall, N. J., Cronin, T. W., & Frank, T. M. (2003). Visual Adaptations in Crustaceans: Chromatic, Developmental, and Temporal Aspects. In S. P. Collin & N. J. Marshall (Eds.), Sensory Processing in Aquatic Environments. (pp. 343‐372). Berlin: Springer‐Verlag. doi: 10.1007/978‐0‐387‐22628‐6_18 18 Visual Adaptations in Crustaceans: Chromatic, Developmental, and Temporal Aspects N. Justin Marshall, Thomas W. Cronin, and Tamara M. Frank Abstract Crustaceans possess a huge variety of body plans and inhabit most regions of Earth, specializing in the aquatic realm. Their diversity of form and living space has resulted in equally diverse eye designs. This chapter reviews the latest state of knowledge in crustacean vision concentrating on three areas: spectral sensitivities, ontogenetic development of spectral sen sitivity, and the temporal properties of photoreceptors from different environments. Visual ecology is a binding element of the chapter and within this framework the astonishing variety of stomatopod (mantis shrimp) spectral sensitivities and the environmental pressures molding them are examined in some detail. The quantity and spectral content of light changes dra matically with depth and water type and, as might be expected, many adaptations in crustacean photoreceptor design are related to this governing environmental factor. Spectral and temporal tuning may be more influenced by bioluminescence in the deep ocean, and the spectral quality of light at dawn and dusk is probably a critical feature in the visual worlds of many shallow-water crustaceans. -

(Callinectes Sapidus) and Stone Crab E-ISSN 2358-2936 (Menippe Mercenaria) Jen L
Nauplius ORIGINAL ARTICLE THE JOURNAL OF THE Grooming behaviors and gill fouling in BRAZILIAN CRUSTACEAN SOCIETY the commercially important blue crab (Callinectes sapidus) and stone crab e-ISSN 2358-2936 www.scielo.br/nau (Menippe mercenaria) www.crustacea.org.br Jen L. Wortham1 orcid.org/0000-0002-4890-5410 Stephanie Pascual1 1 College of Natural and Health Sciences, University of Tampa, 401 West Kennedy Blvd, Tampa, FL, 33606, USA ZOOBANK http://zoobank.org/urn:lsid:zoobank.org:pub:BB965881-F248-4121- 8DCA-26F874E0F12A ABSTRACT Grooming behaviors reduce fouling of body regions. In decapods, grooming time budgets, body regions groomed, and grooming appendages are known in several species; however, little data exists on brachyuran crabs. In this study, grooming behaviors of two commercially important crabs were documented (blue crabs: Callinectes sapidus Rathbun, 1896; stone crabs: Menippe mercenaria Say, 1818). These crabs are harvested by fishermen and knowing their grooming behaviors is valuable, as clean crabs are preferred by consumers and the stone crab fishery consequence of removing one cheliped to grooming behaviors is unknown. Crabs were observed individually and agonistically to determine how grooming behaviors vary in the presence of another conspecific. Both species frequently use their maxillipeds and groom, with the gills being cleaned by epipods. Respiratory and sensory structures were groomed frequently in both species. Removal of a grooming appendage resulted in higher fouling levels in the gills, indicating that grooming behaviors do remove fouling. Overall, stone crabs had a larger individual time budget for grooming, but agonistic grooming time budgets were similar. Stone crab chelipeds are used in grooming, especially cleaning the other cheliped. -

Length-Weight Relationships of Squilla Mantis (Linnaeus, 1758)
International Journal of Fisheries and Aquatic Studies 2018; 6(6): 241-246 E-ISSN: 2347-5129 P-ISSN: 2394-0506 (ICV-Poland) Impact Value: 5.62 Length-weight relationships of Squilla mantis (GIF) Impact Factor: 0.549 IJFAS 2018; 6(6): 241-246 (Linnaeus, 1758) (Crustacea, Stomatopoda, Squillidae) © 2018 IJFAS www.fisheriesjournal.com from Thermaikos Gulf, North-West Aegean Sea, Received: 02-09-2018 Accepted: 03-10-2018 Greece Thodoros E Kampouris (1) Marine Sciences Department, Thodoros E Kampouris, Emmanouil Kouroupakis, Marianna Lazaridou, School of the Environment, and Ioannis E Batjakas University of the Aegean, Mytilene, Lesvos Island, Greece (2) Astrolabe Marine Research, Abstract Mytilene, Lesvos Island, Greece The length-weight relationships (L-W) and the allometric growth profile of the stomatopod Squilla mantis was studied from Thermaikos Gulf, Aegean Sea, Greece. In total, 756 individuals were collected, Emmanouil Kouroupakis by artisanal net fishery and log transformed data were used at the (L-W) relationships assessment. Three (1) Marine Sciences Department, body parts were measured [carapace length (CL), abdominal length (ABL), and telson width (TW)]. Both School of the Environment, females and males demonstrated similarities at their allometric profiles except for CL-W, where females University of the Aegean, present positive allometric profile and males negative. The present findings are mostly in accordance Mytilene, Lesvos Island, Greece (2) Hellenic Centre for Marine with earlier studies from Mediterranean. Research, Institute of Marine Biology, Biotechnology and Keywords: Squilla mantis, stomatopoda, allometric growth, fisheries, Aegean Sea, east Mediterranean Aquaculture, Agios Kosmas, Sea Hellenikon, Athens, Greece 1. Introduction Marianna Lazaridou Squilla mantis (Linnaeus, 1758), spot-tail mantis shrimp, belongs to the Order Stomatopoda, 18, Armpani str. -

The Stalk-Eyed Crustacea of Peru and the Adjacent Coast
\\ ij- ,^y j 1 ^cj^Vibon THE STALK-EYED CRUSTACEA OF PERU AND THE ADJACENT COAST u ¥' A- tX %'<" £ BY MARY J. RATHBUN Assistant Curator, Division of Marine Invertebrates, U. S. National Museur No. 1766.—From the Proceedings of the United States National Museum, '<•: Vol.*38, pages 531-620, with Plates 36-56 * Published October 20, 1910 Washington Government Printing Office 1910 UQS3> THE STALK-EYED CRUSTACEA OF PERU AND THE ADJA CENT COAST. By MARY J. RATHBUN, Assistant Curator, Division of Marine Invertebrates, U. S. National Museum. INTKODUCTION. Among the collections obtained by Dr. Robert E. Coker during his investigations of the fishery resources of Peru during 1906-1908 were a large number of Crustacea, representing 80 species. It was the original intention to publish the reports on the Crustacea under one cover, but as it has not been feasible to complete them at the same time, the accounts of the barnacles a and isopods b have been issued first. There remain the decapods, which comprise the bulk of the collection, the stomatopods, and two species of amphipods. One of these, inhabiting the sea-coast, has been determined by Mr. Alfred O. Walker; the other, from Lake Titicaca, by Miss Ada L. Weckel. See papers immediately following. Throughout this paper, the notes printed in smaller type were con tributed by Doctor Coker. One set of specimens has been returned to the Peruvian Government; the other has been given to the United States National Museum. Economic value.—The west coast of South America supports an unusual number of species of large crabs, which form an important article of food. -

1 NOAA Sea Grant Knauss Marine Policy Fellow NOAA Research
K A I T L Y N L OWDER NOAA Sea Grant Knauss marine policy fellow NOAA Research, Office of International Activities EDUCATION Scripps Institution of Oceanography, University of California, San Diego, La Jolla, CA • Ph.D., Marine Biology with a Specialization in Interdisciplinary Environmental Research, June 2019 • M.S., Biological Oceanography, June 2016 Western Washington University (WWU), Bellingham, WA • B.S., Biology, Marine Emphasis, magna cum laude, June 2014 • B.A., English, Creative Writing, magna cum laude, June 2014 PUBLICATIONS Lowder, K.B., Allen, M.C., Day, J.M., Deheyn, D.D. and Taylor, J.R.A., 2017. Assessment of ocean acidification and warming on the growth, calcification, and biophotonics of a California grass shrimp. ICES Journal of Marine Science, 74(4), pp.1150-1158. Lowder, K.B., Hattingh, R., Day, J.M, and Taylor, J.R.A. Exoskeleton of the California spiny lobster (Panulirus interruptus) is specialized for a multitude of predator defenses. In preparation. Lowder, K.B., deVries, M.S., Hattingh, R., Day, J.M, and Taylor, J.R.A. Exoskeletal predator defenses of the California spiny lobster (Panulirus interruptus) impacted by reduced pH combined with natural pH fluctuations. In preparation. PRESENTATIONS Lowder, K., “Predation preclusion in a changing climate: defenses of California spiny lobster under ocean acidification and warming conditions,” NOAA Inouye Regional Center Science Seminar. Honolulu, HI, Dec 2019. Talk Lowder, K. and J.R.A Taylor. As long as the water’s warm: California spiny lobster larvae get bigger faster in warmer water despite decreases in pH. Association for the Sciences of Limnology and Oceanography. -
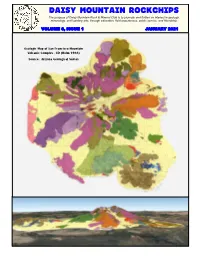
January 2021
Daisy Mountain Rockchips The purpose of Daisy Mountain Rock & Mineral Club is to promote and further an interest in geology, mineralogy, and lapidary arts, through education, field experiences, public service, and friendship. VOLUME 6, ISSUE 1 JANUARY 2021 Geologic Map of San Francisco Mountain Volcanic Complex - 3D (Holm 1988) Source: Arizona Geological Survey Daisy Mountain Rockchips January 2021 2 FOSSILS: PART XIV VOLCANICS OF Kingdom: Animalia Phylum: Arthropoda, NORTHERN ARIZONA Sub-Phylum - Crustacea PART I: San Francisco Peaks - Stratovolcano By Susan Celestian By Susan Celestian In anticipation of a club field trip to visit volcanoes Crustacea includes lobsters, crabs, shrimp, brine of Northern Arizona, around Flagstaff, I am starting shrimp, barnacles, ostracods, crayfish, and a series visiting some of the more interesting and numerous others. They aren’t super common as accessible features. fossils, but they are dominant animals in today’s aquatic environments -- and they sure are tasty! Covering about 1800 square miles, the San Plus, some (such as krill and copepods) are major Francisco Peaks Volcanic Field is home to one players near the bottom of the food chain, where large stratovolcano, several lava domes, and about they feed on plankton, and serve to move those 600 cinder cones. See Figure 1’. nutrients up the chain. Krill are an immensely Activity in the volcanic field began about 6 million important as a food source for whales, seals, and years ago (mya), in the western end of the field, even penguins. And they constitute a large part of and continued progressively east, with the last the overall biomass in the ocean. -

State Finfish Management Project, Southern California Fishery
STATE OF CALIFORNIA-THE RESOURCES AGENCY ARNOLD SCHWARZENEGGER, Governor DEPARTMENT OF FISH AND GAME Marine Region CDFG Monterey Office 20 Lower Ragsdale Drive, Suite 100 Monterey, California 93940 (831) 649-2881 Cruise Report State Finfish Management Project Southern California Fishery-Independent Halibut Trawl Survey Prepared by Sabrina Bell and Travis Tanaka Vessel: F/V Cecelia Dates: February 19-21 and February 28-29, 2008 Location: Halibut trawl grounds off Santa Barbara and Ventura Counties Purpose: Begin an annual monitoring program in southern California that will contribute to the assessment of California halibut and associated species within the halibut trawl grounds. 1. Collect life history data for California halibut, Paralichthys californicus, including length, weight, sex, and otoliths. 2. Tag and release sublegal-sized California halibut (less than 22 in., or 559 mm). 3. Catalog associated incidental species. 4. Record number of halibut damaged, destroyed, or taken because of marine mammal encounters. Procedures: Using Department trawl log data, the existing commercial trawling area was mapped using GIS. The F/V Cecelia was contracted for a 48-hr commitment met by 5 non-consecutive cruise days. Each day the F/V Cecelia embarked the harbor by 0600 and returned by 1700: February 19-21 from the Santa Barbara harbor, and February 28-29 from the Channel Islands Harbor. Most tows were conducted within the halibut trawl grounds using commercial halibut trawl gear with a cod-end mesh of 7.5 in. and were approximately 1 hr in length. Trawl tracks were recorded in real-time using GPS and GIS. During the trawl survey, several considerations were made when choosing the fishing location and course of tows. -

Hermit Crabs - Paguridae and Diogenidae
Identification Guide to Marine Invertebrates of Texas by Brenda Bowling Texas Parks and Wildlife Department April 12, 2019 Version 4 Page 1 Marine Crabs of Texas Mole crab Yellow box crab Giant hermit Surf hermit Lepidopa benedicti Calappa sulcata Petrochirus diogenes Isocheles wurdemanni Family Albuneidae Family Calappidae Family Diogenidae Family Diogenidae Blue-spot hermit Thinstripe hermit Blue land crab Flecked box crab Paguristes hummi Clibanarius vittatus Cardisoma guanhumi Hepatus pudibundus Family Diogenidae Family Diogenidae Family Gecarcinidae Family Hepatidae Calico box crab Puerto Rican sand crab False arrow crab Pink purse crab Hepatus epheliticus Emerita portoricensis Metoporhaphis calcarata Persephona crinita Family Hepatidae Family Hippidae Family Inachidae Family Leucosiidae Mottled purse crab Stone crab Red-jointed fiddler crab Atlantic ghost crab Persephona mediterranea Menippe adina Uca minax Ocypode quadrata Family Leucosiidae Family Menippidae Family Ocypodidae Family Ocypodidae Mudflat fiddler crab Spined fiddler crab Longwrist hermit Flatclaw hermit Uca rapax Uca spinicarpa Pagurus longicarpus Pagurus pollicaris Family Ocypodidae Family Ocypodidae Family Paguridae Family Paguridae Dimpled hermit Brown banded hermit Flatback mud crab Estuarine mud crab Pagurus impressus Pagurus annulipes Eurypanopeus depressus Rithropanopeus harrisii Family Paguridae Family Paguridae Family Panopeidae Family Panopeidae Page 2 Smooth mud crab Gulf grassflat crab Oystershell mud crab Saltmarsh mud crab Hexapanopeus angustifrons Dyspanopeus -

Fiji Fishery Resource Profiles
FIJI FISHERY RESOURCE PROFILES Information for management on 44 of the most important species groups © 2018 Gillett, Preston and Associates ISBN-13: 978–0–9820263–6–6 ISBN-10: 0–9820263–6–6 Cover photo: Keith Ellenbogen Layout and design: Kate Hodge This study was supported by grants from the David and Lucile Packard Foundation and John D. and Catherine T. MacArthur Foundation. Edited by: Sangeeta Mangubhai, Steven Lee, Robert Gillett, Tony Lewis This document should be cited as: Lee, S., A. Lewis, R. Gillett, M. Fox, N. Tuqiri, Y. Sadovy, A. Batibasaga, W. Lalavanua, and E. Lovell. 2018. Fiji Fishery Resource Profiles. Information for Management on 44 of the Most Important Species Groups. Gillett, Preston and Associates and the Wildlife Conservation Society, Suva. 240pp. TABLE OF CONTENTS LIST OF ABBREVIATIONS ................................................................................................8 FOREWORD ........................................................................................................................9 INTRODUCTION ...............................................................................................................10 INVERTEBRATE FISHERY PROFILES ..........................................................................13 1 SEA CUCUMBERS ..............................................................................................................................................14 1.1 The Resource ..................................................................................................................... -

Observations of the Sounds Produced by Swimming in the Spanish
UACE2019 - Conference Proceedings Observations of the Sounds Produced by Swimming in the Spanish Lobster, Scyllarides aequinoctialis (Lund, 1793) By John A. Fornshell * 1 1. U.S. National Museum of Natural History Department of Invertebrate Zoology Smithsonian Institution Washington, D.C. USA *Correspondence: [email protected]; Tel. (571) 426-2398 ABSTRACT The objective of this research project was to study sound production by members of the Scyllaridae, slipper lobsters, specifically the Spanish Lobster Scyllarides aequinoctialis (Lund, 1793) when swimming at or near the surface. This is the first published record of swimming sounds produced by slipper lobsters. Analysis of recordings of swimming sounds produced by S. aequinoctialis in the laboratory indicates a peak of energy in the range of 100 Hz to 1.0 kHz with higher harmonics. The swimming sounds are co-incidental noise produced by slipper lobsters swimming near the surface. Such sounds may have significance for these animals. Spiny lobsters and clawed lobsters produce sounds with a wide range of frequencies and amplitudes. Some of these sounds function as a means of communication with conspecifics. Others produce sounds to ward predators. Incidental sounds produced by feeding, walking or other biological processes may still be significant. Sensory organs that could detect particle motion at short ranges, millimeters to meters, are found on many decapod crustaceans. Key Words: Scyllarides aequinoctialis; of Scyllarides latus; bio-acoustics; noise; INTRODUCTION The ability of spiny lobsters to produce sounds has been well established in the literature for over 134 years. The work of Parker [1] was the first to recognize the existence of two distinct groups of Palinuridae, spiny Lobsters, one producing sounds, the Stridentes, and the other lacking sound producing organs, the Silentes.