This Article Appeared in a Journal Published by Elsevier. the Attached
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Selecting Different Approaches for Palate and Pharynx Surgery
SPECIAL ISSUE 4: INVITED ARTICLE Selecting Different Approaches for Palate and Pharynx Surgery: Palatopharyngeal Arch Staging System Rodolfo Lugo-Saldaña1 , Karina Saldívar-Ponce2 , Irina González-Sáez3 , Daniela Hernández-Sirit4 , Patricia Mireles-García5 ABSTRACT The examination of the anatomical structures involved in the upper airway collapse in patients with the obstructive sleep apnea-hypopnea syndrome (OSAHS) is a key for integrated evaluation of patients. Our proposal is for a noninvasive classification system that guides us about the presence of anatomical differences between the palatopharyngeal muscle (PFM). The functions of the PFM are narrowing the isthmus, descending the palate, and raising the larynx during swallowing; these characteristics give the PFM a special role in the collapse of the lateral pharyngeal wall. Complete knowledge of the anatomy and classification of different variants can guide us to choose the appropriate surgical procedures for the lateral wall collapse. Until now there is not a consensus about description of the trajectory or anatomical variants of the PFM into oropharynx, the distance between both muscles, and the muscle tone. Here we also present the relationship between the lateral wall surgeries currently available (lateral pharyngoplasty by Cahali, expansion sphincteroplasty by Pang, relocation pharyngoplasty by Li, Roman blinds pharyngoplasty by Mantovani, and barbed sutures pharyngoplasty by Vicini) with the proposed classification of the palatopharyngeal arch staging system (PASS). Keywords: -

Absence of Uvula: an Accidental Or an Incidental Finding. J Human Anat
Journal of Human Anatomy ISSN: 2578-5079 Is Uvula Important? Absence of Uvula: An Accidental or an Incidental Finding 1 2 3 4 Vivek J *, Safeer K , Sanjib D and Bhargavi Joshi 1Department of Biochemistry & Basic sciences, Kentucky College of Osteopathic Case Report Volume 3 Issue 2 Medicine, USA Received Date: September 12, 2019 2Department of Anatomy & Embryology, Windsor University School of Published Date: October 21, 2019 Medicine, Saint Kitts and Nevis DOI: 10.23880/jhua-16000142 3Department of Pharmacology, Govt Medical College, Ratlam, India 4Research Volunteer, Windsor University School of Medicine, St Kitts and Nevis *Corresponding author: Vivek Joshi, MD, Associate Professor Biochemistry, Department of Basic Science, Kentucky College of Osteopathic Medicine, 147 Sycamore Street, Hambley Blvd, University of Pikeville (UPike), Pikeville, KY, 41501, USA, Tel : 606-218-5552; Email: [email protected] Abstract Introduction: Absence of the uvula is very rare in the general population, which is mostly acquired secondary to surgery or is rarely congenitally absent since birth. Uvula is a small band of connective tissue, gland and small muscle fibers and is documented to be useful in speech, lubrication and central support of the palatopharyngeal arch during swallowing. Cultural practice of uvulectomy is very common in African countries as a treatment or prophylactic measure for chronic cough or frequent respiratory infection. Congenital absence of uvula is a rare condition and is also accompanied by other genetic abnormalities such as cleft lip or cleft palate. Case Report: This case report is based on an accidental finding in a 20-year-old African-American male who was acting as a standardized patient in a clinical course at a medical college. -

General Anatomy of Gastro-Intestinal System
General Anatomy of Gastro-IntesTinal System The teeth, Oral cavity, Tongue, Salivary glands, Pharynx. Their vessels and innervation IKIvo Klepáček Primordium of the alimentary canal (GastroInTestinal Canal) GIT devel– systema gastropulmonale – it develops from the embryonal intestine (entoderm) ; lower respiratory structurses are splitted from intewstine as a tracheobronchial pouch Ventral (head) intestine part is added to ectodermal pouch called stomodeum, caudal part of the intestine is added to ectodermal pouch called proctodeum Division of the alimentary tract: 1) oral ectodermal segment 2) main entodermal segment 3) caudal ectodermal segment děivision of the main segment: ventral gut (foregut – to biliary duct opening) middle gut (midgut – to 2/3 colon) IKdorsal gut (hindgut – to upper part of the anal canal Digestive System: Oral cavity (ectodermal origin) The gut and ist derivatives (entodermal origin) is devided in four sections: 1. Pharyngeal gut or pharynx 2. Foregut - esophagus, stomach, ¼ of duodenum, liver and gallblader, pancreas 3. Midgut – ¾ of duodenum, jejujnum, ilium, colon caecum, colon ascendens and 2/3 of colon transversum 4. Hindgut – 1/3 of colon transversum, colon descendens, colon sigmoideum, colon rectum, IKcanalis analis IK Alimentary tube (canal) - general structure – tunica mucosa (mucous membrane 1 • epithelium • lamina propria mucosae (lymph tissue) • lamina muscularis mucosae – tunica submucosa (submucous layer) – vessels, erves (plexus submucosus Meissneri) – tunica muscularis externa 7 (outer -

A Decision Rule for Diagnostic Testing in Obstructive Sleep Apnea
A Decision Rule for Diagnostic Testing in Obstructive Sleep Apnea Willis H. Tsai, John E. Remmers, Rollin Brant, W. Ward Flemons, Jan Davies, and Colin Macarthur Department of Medicine, Division of Respiratory Medicine; Department of Community Health Sciences; and Department of Anesthesia, University of Calgary, Calgary, AB, Canada Obstructive sleep apnea (OSA) is traditionally diagnosed using over- hourϪ1 or more) of 5.17% and 81%, respectively. In contrast, night polysomnography. Decision rules may provide an alternative patients with the lowest clinical score had a likelihood ratio to polysomnography. A consecutive series of patients referred to of 0.25 and a post-test probability of OSA of 17%. a tertiary sleep center underwent prospective evaluation with the A morphometric model developed by Kushida and col- upper airway physical examination protocol, followed by determi- leagues had an OSA diagnostic sensitivity and specificity of nation of the respiratory disturbance index using a portable moni- 98% and 100%, respectively; however, selection bias was a tor. Seventy-five patients were evaluated with the upper airway physical examination protocol. Historic predictors included age, potential concern (3). Nevertheless, the model illustrated the snoring, witnessed apneas, and hypertension. Physical examination– potential value of physical examination–based decision rules based predictors included body mass index, neck circumference, in clinical decision-making. mandibular protrusion, thyro–rami distance, sterno–mental distance, Current decision rules have only intermediate diagnostic sterno–mental displacement, thyro–mental displacement, cricomen- characteristics and are frequently too cumbersome, either tal space, pharyngeal grade, Sampsoon-Young classification, and over- arithmetically or logistically, for bedside implementation (2, bite. A decision rule was developed using three predictors: a crico- 4–10). -
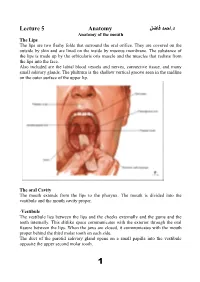
Lecture 5 Anatomy احمد فاضل د
د.احمد فاضل Lecture 5 Anatomy Anatomy of the mouth The Lips The lips are two fleshy folds that surround the oral orifice. They are covered on the outside by skin and are lined on the inside by mucous membrane. The substance of the lips is made up by the orbicularis oris muscle and the muscles that radiate from the lips into the face. Also included are the labial blood vessels and nerves, connective tissue, and many small salivary glands. The philtrum is the shallow vertical groove seen in the midline on the outer surface of the upper lip. The oral Cavity The mouth extends from the lips to the pharynx. The mouth is divided into the vestibule and the mouth cavity proper. -Vestibule The vestibule lies between the lips and the cheeks externally and the gums and the teeth internally. This slitlike space communicates with the exterior through the oral fissure between the lips. When the jaws are closed, it communicates with the mouth proper behind the third molar tooth on each side. The duct of the parotid salivary gland opens on a small papilla into the vestibule opposite the upper second molar tooth. 1 -Mouth Proper The mouth proper has a roof and a floor. Roof of Mouth The roof of the mouth is formed by the hard palate in front and the soft palate behind. Floor of Mouth The submandibular duct of the submandibular gland opens onto the floor of the mouth on the summit of a small papilla on either side of the frenulum of the tongue. -

The Mouth the Mouth Extends from the Lips to the Oropharyngeal Isthmus, That Is, the Junction of the Mouth with the Pharynx
The Mouth The mouth extends from the lips to the oropharyngeal isthmus, that is, the junction of the mouth with the pharynx. It is subdivided into the vestibule, which lies between the lips and cheek externally and the gums and teeth internally, and the mouth cavity proper, which lies within the alveolar arches, gums, and teeth. The vestibule is a slitlike space that communicates with the exterior through the oral fissures. When the jaws are closed, it communicates with the mouth cavity proper behind the third molar tooth on each side. Superiorly and inferiorly, the vestibule is limited by the reflection of the mucous membrane from the lips and cheeks onto the gums. The cheek forms the lateral wall of the vestibule and is made up of the buccinator muscle, which is covered on the outside by fascia and skin and is lined by mucous membrane. Opposite the upper second molar teeth, a small papilla is present on the mucous membrane, marking the opening of the duct of the parotid salivary gland. The mouth proper has a roof, which is formed by the hard palate in front and the soft palate behind. The floor is formed by the anterior two-thirds of the tongue and by the reflection of the mucous membrane from the sides of the tongue to the gum on the mandible. In the midline, a fold of mucous membrane called frenulum of the tongue connects the undersurface of the tongue to the floor of the mouth. On each side of the frenulum is a small papilla, on the summit of which is the orifice of the duct of the submandibular salivary gland. -
Clinical Guidelines for Treatment of Impalement Injuries of the Oropharynx in Children
clinical section Clinical guidelines for treatment of impalement injuries of the oropharynx in children Ari Kupietzky, DMD, MSc Dr. Kupietzky is in private practice, Jerusalem, Israel, and a Diplomate of The American Board of Pediatric Dentistry. rodental injury in children is a cause of much concern and anxiety for the patient and parent. Most pediat- Oric dentists are well-acquainted and capable of treat- ment of trauma to the teeth and their supporting structures. However, the anatomical area posterior to the hard palate is less familiar to the pediatric dentist. The active toddler and young child are likely to turn first to their familiar dentist if and when they experience an impalement injury. The pediat- ric dentist needs to be in a position to provide proper emergency treatment and reassurance in such cases. The objectives of the present report are to describe an im- palement injury in a 6-year old, review the relevant anatomy, modes of treatment, and possible sequela associated with such injuries. Mini review of terminology, prevalence and etiology Impalement injuries of the oropharynx and particularly of the soft palate are relatively common in children.1-5 Impalement injuries of the soft palate have been reported in patients of all Fig 1. Landmark anatomy of oropharynx and soft palate region: 1. Palatine ages, ranging from the newborn to geriatric patients. However, tonsil. 2. Supratonsillar fossa. 3. Mid soft palate. 4. Hard palate. 5. Palatopharyngeal arch. 6. Interior pillar. The clinician should be able to young children, particularly toddlers, are most involved in these describe tha area of injury. -
Oral Cavity, Tongue & Palate
By Prof Nabil A Hasan Head of Anatomy Department Lies in the roof of the oral cavity Has two parts: hard • Hard (bony) palate soft palate anteriorly • Soft (muscular) palate posteriorly Lies in the roof of the oral cavity Forms the floor of the nasal cavity Formed by: • Palatine processes of maxillae in front • Horizontal plates of palatine bones behind Bounded by alveolar arches Posteriorly, continuous with soft palate Its undersurface covered by mucoperiosteum Shows transverse ridges in the anterior parts Attached to the posterior border of the hard palate Covered on its upper and lower surfaces by mucous membrane Composed of: • Muscle fibers • An aponeurosis • Lymphoid tissue • Glands • Blood vessels • Nerves Fibrous sheath Attached to posterior border of hard palate Is expanded tendon of tensor velli palatini Splits to enclose musculus uvulae Gives origin & insertion to palatine muscles Tensor veli palatini • Origin: Scaphoid fossa pterygoid bone & auditory tube • Insertion: superior surface of palatine aponeurosis • Action: Tenses soft palate Levator veli palatini • Origin:petrous part of the temporal bone & auditory tube • Insertion: superior surface of palatine aponeurosis • Action: Raises soft palate to close the posterior pharyngeal wall Musculus uvulae Origin: posterior border of hard palate Insertion: mucosa of uvula Action: Elevates uvula and contact the posterior pharyngeal wall Palatoglossus • Origin: inferior surface of palatine aponeurosis • Insertion: side of tongue • Action: pulls root of tongue upward, -
Digestive System
Chapter 25 *Lecture PowerPoint The Digestive System *See separate FlexArt PowerPoint slides for all figures and tables preinserted into PowerPoint without notes. Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Introduction • Most nutrients we eat cannot be used in existing form – Must be broken down into smaller components before the body can make use of them • Digestive system—essentially a disassembly line – To break down nutrients into a form that can be used by the body – To absorb them so they can be distributed to the tissues • Gastroenterology—the study of the digestive tract and the diagnosis and treatment of its disorders 25-2 General Anatomy and Digestive Processes • Expected Learning Outcomes – List the functions and major physiological processes of the digestive system. – Distinguish between mechanical and chemical digestion. – Describe the basic chemical process underlying all chemical digestion, and name the major substrates and products of this process. 25-3 General Anatomy and Digestive Processes Cont. – List the regions of the digestive tract and the accessory organs of the digestive system. – Identify the layers of the digestive tract and describe its relationship to the peritoneum. – Describe the general neural and chemical controls over digestive function. 25-4 Digestive Function • Digestive system—the organ system that processes food, extracts nutrients from it, and eliminates the residue 25-5 Digestive Function • Five stages of digestion – Ingestion: selective intake of -
Digestive-System-1.1.Pdf
Digestive System By: Donna Browne Digestive System By: Donna Browne Online: < http://legacy.cnx.org/content/col11761/1.1/ > OpenStax-CNX This selection and arrangement of content as a collection is copyrighted by Donna Browne. It is licensed under the Creative Commons Attribution License 4.0 (http://creativecommons.org/licenses/by/4.0/). Collection structure revised: February 23, 2015 PDF generated: February 23, 2015 For copyright and attribution information for the modules contained in this collection, see p. 72. Table of Contents 1 Digestive System Module 1: Overview of the Digestive System ..............................1 2 Digestive System Module 2: Processes and Regulation .......................................7 3 Digestive System Module 3: The Mouth, Pharynx, and Esophagus ........................13 4 Digestive System Module 4: The Stomach ....................................................27 5 Digestive System Module 5: The Small and Large Intestines ...............................33 6 Digestive System Module 6: Accessory Organs in Digestion: The Liver, Pancreas, and Gallbladder ...................................................................43 7 Digestive System Module 7: Chemical Digestion and Absorption: A Closer Look ............................................................................................51 Glossary .............................................................................................61 Index ................................................................................................68 Attributions -

Influence of the Palatine Tonsil Grade on the Morphology of the Maxillary and Mandibular Dental Arches
Int. J. Morphol., 38(5):1201-1207, 2020. Influence of the Palatine Tonsil Grade on the Morphology of the Maxillary and Mandibular Dental Arches Influencia del Grado de las Tonsilas Palatinas en la Morfología de los Arcos Dentales Maxilar y Mandibular Inés Pérez1; Nilton Alves2,3; Claudia Lizana4 & Naira Figueiredo Deana5,6 PÉREZ, I.; ALVES, N.; LIZANA, C. & DEANA, N. F. Influence of the palatine tonsil grade on the morphology of the maxillary and mandibular dental arches. Int. J. Morphol., 38(5):1201-1207, 2020. SUMMARY: Enlarged palatine tonsils and adenoids are thought to cause obstruction of the upper airway, triggering changes in breathing patterns, which in turn lead to dentofacial alterations, including malocclusions. The object of the present study was to correlate the size (grade) of the palatine tonsil with measurements of the maxillary and mandibular dental arches in children. This was an observational cross-sectional study carried out in 35 children aged between 6 and 11 years. The inter- and intra-arch parameters were measured (horizontal, vertical and sagittal analyses) by making plastercasts and then taking measurements with callipers. The tonsil size was classified in 5 grades from 0-4, using the Brodsky tonsil grading scale. The researcher was calibrated prior to carrying out the measurements. Pearson's chi-squared test was used and Pearson's correlation coefficient was calculated. The SPSS v.22.0 software was used, with a significance threshold of 5 %. Six patients were classified as grade 1, sixteen grade 2, eleven grade 3, two grade 4 and none of the patients presented grade 0. -

Tongue and Salivary Glands
Tongue and Salivary Glands 1. Tongue 2. Salivary glands – minor and major salivary glands 3. Isthmus of the fauces 4. Tonsils SPLANCHNOLOGY Tongue . Tongue – lingua (Gr. glossa): on the floor of the mouth a muscular organ possessing great mobility important role in digestion: active role in chewing (mastication) involved in sucking and deglutition reflex essential role in articulation and phonation the primary organ of taste Prof. Dr. Nikolai Lazarov 2 SPLANCHNOLOGY Phylogenesis of the tongue . Phylogenesis: fishes a fold in the floor of the mouth epithelial lining, no glands and muscles connective tissue of hyoid arch amphibians and reptiles tactile organ well-developed mucous glands intrinsic muscles mammals important organ of the digestive system; vital role in feeding and sucking highly differentiated mucosa proper musculature Prof. Dr. Nikolai Lazarov 3 SPLANCHNOLOGY Embryonic development . Onthogenesis: begin – 4 mo. origin – from branchial arches body of tongue – Ist pharyngeal arch: mesodermal cells from the 1st (mandibular) pharyngeal arch two lateral lingual swellings – 5 we. median lingual swelling, tuberculum impar root of tongue: hypobranchial eminence, copula of His – II-IV pharyngeal arch mucosa: body – 1st pharyngeal arch root – 3rd-4th arches tongue musculature: from occipital (postotic) myotomes (3rd and 4th occipital myotomes) taste buds: 7 we. Prof. Dr. Nikolai Lazarov 4 SPLANCHNOLOGY Structure of the tongue . Parts of the tongue: body, corpus linguae tip, apex linguae root, radix linguae divided by a V-shaped furrow, sulcus terminalis . Surface anatomy: dorsal surface, dorsum linguae oral (presulcal) part (pars oralis) pharyngeal (postsulcal) part (pars pharyngealis) sulcus terminalis foramen cecum median furrow, sulcus medianus inferior surface, facies inferior plica fimbriata frenulum linguae caruncula sublingualis sublingual fold margin of the tongue, margo linguae Prof.