Proceedings of the United States National Museum
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

The Chiton Radula: a Unique Model for Biomineralization Studies
4 The Chiton Radula: A Unique Model for Biomineralization Studies Lesley R. Brooker1 and Jeremy A. Shaw2 1University of the Sunshine Coast 2Centre for Microscopy, Characterisation & Analysis University of Western Australia Australia 1. Introduction Over the course of evolution, a range of strategies have been developed by different organisms to produce unique materials and structures perfected for their specific function. This biological mastery of materials production has inspired the birth of the new discipline of biomaterials through biomimicry (Birchall, 1989). Chitons (Mollusca: Polyplacophora) are slow moving, bilaterally symmetrical and dorso- ventrally flattened molluscs that are commonly found on hard substrata in intertidal regions of coastlines around the world (Kaas & Jones, 1998). All species are characterized by a series of eight dorsal, articulating shell plates or valves, which may be embedded, to varying degrees, in a fleshy, muscular girdle (Kaas & Jones, 1998) (Figure 1). Approximately 750 living species are known, and while intertidal regions are home to the majority of chitons, a number of species can be found at depths of up to 8000m where they feed on detrital material (Kaas & Jones, 1998). Fig. 1. Photograph of the dorsal surface of the chiton Acanthopleura gaimardi, showing the eight overlapping aragonite plates surrounded by the fleshy girdle, which, in this species, is covered in small aragonite spines. Chitons feed by rasping macro- and micro-algae from the rocks on which they live through the use of a radula. The radula has been coined as a conveyor belt of continuously developing www.intechopen.com 66 Advanced Topics in Biomineralization teeth, replaced by new teeth as they are worn and lost. -

The Recent Molluscan Marine Fauna of the Islas Galápagos
THE FESTIVUS ISSN 0738-9388 A publication of the San Diego Shell Club Volume XXIX December 4, 1997 Supplement The Recent Molluscan Marine Fauna of the Islas Galapagos Kirstie L. Kaiser Vol. XXIX: Supplement THE FESTIVUS Page i THE RECENT MOLLUSCAN MARINE FAUNA OF THE ISLAS GALApAGOS KIRSTIE L. KAISER Museum Associate, Los Angeles County Museum of Natural History, Los Angeles, California 90007, USA 4 December 1997 SiL jo Cover: Adapted from a painting by John Chancellor - H.M.S. Beagle in the Galapagos. “This reproduction is gifi from a Fine Art Limited Edition published by Alexander Gallery Publications Limited, Bristol, England.” Anon, QU Lf a - ‘S” / ^ ^ 1 Vol. XXIX Supplement THE FESTIVUS Page iii TABLE OF CONTENTS INTRODUCTION 1 MATERIALS AND METHODS 1 DISCUSSION 2 RESULTS 2 Table 1: Deep-Water Species 3 Table 2: Additions to the verified species list of Finet (1994b) 4 Table 3: Species listed as endemic by Finet (1994b) which are no longer restricted to the Galapagos .... 6 Table 4: Summary of annotated checklist of Galapagan mollusks 6 ACKNOWLEDGMENTS 6 LITERATURE CITED 7 APPENDIX 1: ANNOTATED CHECKLIST OF GALAPAGAN MOLLUSKS 17 APPENDIX 2: REJECTED SPECIES 47 INDEX TO TAXA 57 Vol. XXIX: Supplement THE FESTIVUS Page 1 THE RECENT MOLLUSCAN MARINE EAUNA OE THE ISLAS GALAPAGOS KIRSTIE L. KAISER' Museum Associate, Los Angeles County Museum of Natural History, Los Angeles, California 90007, USA Introduction marine mollusks (Appendix 2). The first list includes The marine mollusks of the Galapagos are of additional earlier citations, recent reported citings, interest to those who study eastern Pacific mollusks, taxonomic changes and confirmations of 31 species particularly because the Archipelago is far enough from previously listed as doubtful. -
Atlanta Ariejansseni, a New Species of Shelled Heteropod from the Southern Subtropical Convergence Zone (Gastropoda, Pterotracheoidea)
A peer-reviewed open-access journal ZooKeys 604: 13–30 (2016) Atlanta ariejansseni, a new species of shelled heteropod.... 13 doi: 10.3897/zookeys.604.8976 RESEARCH ARTICLE http://zookeys.pensoft.net Launched to accelerate biodiversity research Atlanta ariejansseni, a new species of shelled heteropod from the Southern Subtropical Convergence Zone (Gastropoda, Pterotracheoidea) Deborah Wall-Palmer1,2, Alice K. Burridge2,3, Katja T.C.A. Peijnenburg2,3 1 School of Geography, Earth and Environmental Sciences, Plymouth University, Drake Circus, Plymouth, PL4 8AA, UK 2 Naturalis Biodiversity Center, Darwinweg 2, 2333 CR Leiden, The Netherlands3 Institute for Biodiversity and Ecosystem Dynamics (IBED), University of Amsterdam, P. O. Box 94248, 1090 GE Amster- dam, The Netherlands Corresponding author: Deborah Wall-Palmer ([email protected]) Academic editor: N. Yonow | Received 21 April 2016 | Accepted 22 June 2016 | Published 11 July 2016 http://zoobank.org/09E534C5-589D-409E-836B-CF64A069939D Citation: Wall-Palmer D, Burridge AK, Peijnenburg KTCA (2016) Atlanta ariejansseni, a new species of shelled heteropod from the Southern Subtropical Convergence Zone (Gastropoda, Pterotracheoidea). ZooKeys 604: 13–30. doi: 10.3897/zookeys.604.8976 Abstract The Atlantidae (shelled heteropods) is a family of microscopic aragonite shelled holoplanktonic gastro- pods with a wide biogeographical distribution in tropical, sub-tropical and temperate waters. The arago- nite shell and surface ocean habitat of the atlantids makes them particularly susceptible to ocean acidifica- tion and ocean warming, and atlantids are likely to be useful indicators of these changes. However, we still lack fundamental information on their taxonomy and biogeography, which is essential for monitoring the effects of a changing ocean. -

DISCUSSION of the FLORA of GUADALUPE ISLAND Dr. Reid Moran1
DISCUSSION OF THE FLORA OF GUADALUPE ISLAND Dr. Reid Moran1: Guadalupe Island lies about 250 miles south- southwest of San Diego, California, and about 160 miles off the peninsula of Baja California, Mexico. Volcanic in origin and sep arated from the peninsula by depths of about 12,000 feet, it is clearly an oceanic island. Among the vascular plants recorded from Guadalupe Island and its islets, apparently 164 species are native. Goats, introduced more than a century ago, have eliminated some species and re duced others nearly to extinction. Outer Islet, a goatless refugium two miles south of the main island, has a native florula of 36 species. Nine of these (including Euphorbia misera and Lavatera occidentalis) are very scarce on the main island, largely confined to cliffs inaccessible to goats; another one (Coreopsis gigantea) has not been collected there since 1875; and five others (includ ing Lavatera lindsayi, Dudleya guadalupensis, and Rhus integri¬ folia) have never been recorded from there. Although it is not known that these five ever did occur on the main island, presum ably they did but were exterminated by the goats. These five, comprising 14 per cent of the native florula of Outer Islet, give the only suggestion we have as to how many species must have been eliminated from the main island by the goats before they could be found by botanists. Also reported from Guadalupe Island are 42 species that prob ably are not native. Several of these, each found only once, ap parently have not persisted; but, with the severe reduction of many native plants by the goats, other introduced plants have become abundant. -

FONDO INVESTIGACION PESQUERA Bases Biológicas ... -.:: CESSO
INFORME FINAL CORREGIDO FIP Nº 2002-16 Bases biológicas para la evaluación y manejo de metapoblaciones de loco en la III y IV Regiones • Abril, 2005 • REQUIRENTE FONDO DE INVESTIGACIÓN PESQUERA, FIP Presidente del Consejo: Felipe Sandoval Precht EJECUTOR INSTITUTO DE FOMENTO PESQUERO, IFOP Jefe División Investigación Pesquera: Mauricio Braun Alegría Director Ejecutivo: Guillermo Moreno Paredes • Abril, 2005 • JEFE DE PROYECTO JORGE GONZÁLEZ Y. AUTORES JORGE GONZÁLEZ CARLOS TAPIA ALVARO WILSON WOLFGANG STOTZ JOSÉ MARÍA (LOBO) ORENSANZ ANA PARMA JUAN VALERO MÓNICA CATRILAO JORGE GARRIDO COLABORADORES CARLOS CORTÉS CÉSAR GUEVARA CRISTIAN CANALES DOMINGO LANCELLOTTI JAIME ABURTO MARCELO VALDEBENITO LUIS CAILLAUX • Abril, 2005 • INSTITUTO DE FOMENTO PESQUERO / DIVISIÓN INVESTIGACIÓN PESQUERA RESUMEN EJECUTIVO En la III y IV Regiones las áreas de manejo y explotación de recursos bentónicos (AMERB) se han establecido como la principal herramienta de manejo del recurso loco (Concholepas concholepas). Sin embargo, su aplicabilidad para la administración y aporte en términos del desarrollo sustentable, aunque sugerentes, requieren de una evaluación objetiva, en cuanto al manejo de carácter discreto que se ha establecido para cada AMERB, desconociendo la eventual interacción espacial de las mismas. En este marco, se desarrolla una aproximación para evaluar el desempeño de las AMERBs desde una perspectiva integral bajo un enfoque metapoblacional. Si bien, el estudio está orientado a proporcionar la información de campo que permita probar la hipótesis de metapoblaciones en el recurso loco en la III y IV Regiones y proponer un modelo inicial que incorpore las tasas de extinción y colonización de los parches o poblaciones locales. Se debe entender que para el caso del recurso loco, la metapoblación no es un conjunto de “parches” discretos, (concepto asociado al modelo original de Levins), sino a un continuo heterogéneo conformado por zonas de productividad diferencial. -

Type Material of Adelomelon Indigestus Von Ihering, 1908 (Gastropoda, Volutidae)
Short communication Type material of Adelomelon indigestus von Ihering, 1908 (Gastropoda, Volutidae) Fabio Wiggers1* Inga Ludmila Veitenheimer-Mendes2 Universidade Federal do Rio Grande do Sul, Programa de Pós-Graduação em Biologia Animal Av. Bento Gonçalves, 9500, Porto Alegre, CEP 91501-970 Rio Grande do Sul, Brazil *Autor para correspondência [email protected] 2 [email protected] Submetido em 21/02/2005 Aceito para publicação em 28/04/2005 Resumo Material tipo de Adelomelon indigestus von Ihering, 1908 (Gastropoda, Volutidae). Adelomelon indigestus von Ihering, 1908 é considerado um sinônimo de Adelomelon beckii (Broderip, 1836), entretanto a categoria taxonômica de seu material tipo não estava claro. Após a localização e exame dos dois exemplares citados na descrição original de A. indigestus os mesmos foram reconhecidos como síntipos. De acordo com o Código Internacional de Nomenclatura Zoológica, não havendo designação de lectótipo, a localidade-tipo deve incluir tanto Guaratiba, Rio de Janeiro quanto São Sebastião, São Paulo, ambos localizados no litoral sudeste do Brasil. Unitermos: Taxonomia, material tipo, Volutidae, Adelomelon indigestus, Oeste do Atlântico Sul Biotemas, 18 (2): 227 - 231, 2005 F. Wiggers e I. L. Veitenheimer-Mendes Abstract Adelomelon indigestus von Ihering, 1908 is known to be a synonym of Adelomelon beckii (Broderip, 1836) but the taxonomic status of its type material was not previously clear. The specimens cited in the original description of A. indigestus were located and recognized as syntypes. Under the rule of the International Code of Zoological Nomenclature, the type locality is proposed to include Guaratiba (Rio de Janeiro) and São Sebastião (São Paulo) both on the southeastern littoral of Brazil. -

Distribution Patterns of Pelagic Gastropods at the Cape Verde Islands Holger Ossenbrügger
Distribution patterns of pelagic gastropods at the Cape Verde Islands Holger Ossenbrügger* Semester thesis 2010 *GEOMAR | Helmholtz Centre for Ocean Research Kiel Marine Ecology | Evolutionary Ecology of Marine Fishes Düsternbrooker Weg 20 | 24105 Kiel | Germany Contact: [email protected] Contents 1. Introduction . .2 1.1. Pteropods . 2 1.2. Heteropods . 3 1.3. Hydrography . 4 2. Material and Methods . 5 3. Results and Discussion . 7 3.1. Pteropods . 7 3.1.1. Species Composition . 7 3.1.2. Spatial Density Distribution near Senghor Seamount . .. 9 3.1.3. Diel Vertical Migration . 11 3.2. Heteropods . 17 3.2.1. Species Composition . .17 3.2.2. Spatial Density Distribution near Senghor Seamount . .17 3.2.3. Diel Vertical Migration . 18 4. Summary and directions for future research . 19 References . 20 Acknowledgements . 21 Attachment . .22 1. Introduction 1.1. Pteropods Pteropods belong to the phylum of the Mollusca. They are part of the class Gastropoda and located in the order Ophistobranchia. The pteropods are divided into the orders Thecosomata and Gymnosomata. They are small to medium sized animals, ranging from little more than 1mm for example in many members of the Genus Limacina to larger species such as Cymbulia peroni, which reaches a pseudoconch length of 65mm. The mostly shell bearing Thecosomata are known from about 74 recent species worldwide and are divided into five families. The Limacinidae are small gastropods with a sinistrally coiled shell; they can completely retract their body into the shell. Seven recent species of the genus Limacina are known. The Cavoliniidae is the largest of the thecosomate families with about 47 species with quite unusually formed shells. -

Pyramidellidae Turbonilla
Pyramidellidae Subfamily Turbonillinae Genus Turbonilla Species recorded from the SCB By Tony Phillips DCE At the October 2010 SCAMIT meeting it was suggested by Pat LaFollette for us to take high quality, detailed photographs of our Turbonilla, Pyrgiscus and Odostomia species (don’t forget scale bars, locality and depth information!) and share with him and with each other. If it is possible to assign it unequivocally to a described species, he will aid us in giving specimens names…... He also suggested we use Abbott 1974 as a primary resource..... Subgenera of Turbonilla (Oldroyd 1927 & Abbot 1974) • Bartschella Iredale 1917 - valid genus • Chemnitzia d’Orbigny 1840 - Turbonilla • Mormula A. Adams 1863 - valid genus, not in subfamily Turbonillinae • Pyrgiscus Philippi 1841 - valid genus • Pyrgolampros Sacco 1892 - Turbonilla • Strioturbonilla Sacco 1892 – valid genus, not in subfamily Turbonillinae • Turbonilla Risso 1826 – valid genus * WoRMS 2016 for subfamily Turbonillinae Genera listed under subfamily Turbonillinae (according to WoRMS 2016) Asmunda Dall & Bartsch 1904 Kleinella A. Adams 1860 Bacteridella Saurin 1959 Nisipyrgiscus Robba 2013 Bartschella Iredale 1916 Nisiturris Dall & Bartsch 1906 Bouchetmella Penas & Rolan 2016 Puposyrnola Cossmann 1921 Careliopsis Morch 1875 Pyrgiscus Philippi 1841 Colsyrnola Iredale 1929 Pyrgolidium Monterosato 1884 Ebalina Thiele 1929 Pyrgostylus Monterosato 1884 Eulimella Forbes & M’Andrew 1846 Raoulostraca Oliver 1915 Exesilla Laseron 1959 Syrnola A. Adams 1860 Gispyrella Laws 1937 Turbolidium Robba 2013 Hamarilla Eames & Wilkins 1959 Turbonilla Risso 1826 Iphiana Dall & Bartsch 1904 Considering the current state of uncertainty of the suprageneric classification, we have preferred not to add to the confusion, and have kept our work at the specific level. Therefore we shall use only the genus name Turbonilla for all the species included….(Penas & Rolan, 2010) ….as most of the turbonillids, the majority of species cannot yet be defensibly seated in an appropriate genus. -

The Conservation of Forest Genetic Resources Case Histories from Canada, Mexico, and the United States
The Conservation of Forest Genetic Resources Case Histories from Canada, Mexico, and the United States value of gene banks in the conservation of forest genetic resources. By E Thomas Ledig, J.JesusVargas-Hernandez, and Kurt H. Johnsen Prepared as a task of the Forest Genetic Resources Study Group/North American Forestry CornrnissioniFood and Agriculture Organization of the United Nations. Reprinted from the Joul-rzal of Forestry, Vol. 96, No. 1, January 1998. Not for further reproduction. e Conservation of Forest Case Histories from Canada, Mexico, and the United States he genetic codes of living organ- taken for granted: release of oxygen and isms are natural resources no less storage of carbon, amelioration of cli- T than soil, air, and water. Genetic mate, protection of watersheds, and resources-from nucleotide sequences others. Should genetic resources be lost, in DNA to selected genotypes, popula- ecosystem function may also be dam- tions, and species-are the raw mater- aged, usually expressed as a loss of pri- ial in forestry: for breeders, for the for- mary productivity, the rate at which a est manager who produces an eco- plant community stores energy and pro- nomic crop, for society that reaps the duces organic matter (e.g., Fetcher and environmental benefits provided by Shaver 1990). Losses in primary pro- forests, and for the continued evolu- ductivity result in changes in nutrient tion of the species itself. and gas cycling in Breeding, of course, The loss g~f;a ecosystems (Bormann requires genetic variation. and Likens 1979). Continued improvement p population is Genetic diversity is in medicines, agricultural the most basic element By F. -

Phylogeny of the Sea Hares in the Aplysia Clade Based on Mitochondrial DNA Sequence Data
Lawrence Berkeley National Laboratory Lawrence Berkeley National Laboratory Title Phylogeny of the sea hares in the aplysia clade based on mitochondrial DNA sequence data Permalink https://escholarship.org/uc/item/0fv9g804 Authors Medina, Monica Collins, Timothy Walsh, Patrick J. Publication Date 2004-02-20 Peer reviewed eScholarship.org Powered by the California Digital Library University of California PHYLOGENY OF THE SEA HARES IN THE APLYSIA CLADE BASED ON 1,2 MITOCHONDRIAL DNA SEQUENCE DATA MÓNICA MEDINA , TIMOTHY 3 1 COLLINS , AND PATRICK J. WALSH 1 Rosenstiel School of Marine and Atmospheric Science, Division of Marine Biology and Fisheries, University of Miami, 4600 Rickenbacker Causeway, Miami, FL 33149 USA. RUNNING HEAD: Aplysia mitochondrial phylogeny 2 present address: Joint Genome Institute, 2800 Mitchell Drive B400, Walnut Creek, CA 94598 e-mail: [email protected] phone: (925)296-5633 fax: (925)296-5666 3 Department of Biological Sciences, Florida International University, University Park, Miami, FL 33199 USA ABSTRACT Sea hare species within the Aplysia clade are distributed worldwide. Their phylogenetic and biogeographic relationships are, however, still poorly known. New molecular evidence is presented from a portion of the mitochondrial cytochrome oxidase c subunit 1 gene (cox1) that improves our understanding of the phylogeny of the group. Based on these data a preliminary discussion of the present distribution of sea hares in a biogeographic context is put forward. Our findings are consistent with only some aspects of the current taxonomy and nomenclatural changes are proposed. The first, is the use of a rank free classification for the different Aplysia clades and subclades as opposed to previously used genus and subgenus affiliations. -
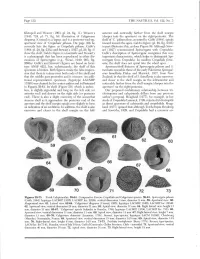
(1943: 724, Pi. 71, Fig. 16) Illustration of Calyptraea (Deeper Into the Aperture) on the Right/Posterior
Page 118 THE NAUTILUS, Vol. 122, No. 3 Kleinpeil and Weaver (1963, pi. 24, fig. 11). Weavers anterior and noticeably farther from the shell margin (1943: 724, pi. 71, fig. 16) illustration of Calyptraea (deeper into the aperture) on the right/posterior. The diegoana (Conrad) is a lapsus and is a posterior-end-up, shelf of 'C.y pileum thus, as noted by Gabb (1864), spirals apertural view of 'Crepidula' pileum. On page 356 he inward toward the apex. Gabb's figure (pi. 29, fig. 233b) correctly lists the figure as Crepidula pileum. Gabb's in part illustrates this, as does Figure 59. Although Stew- (1864: pi. 29, fig. 233a) and Stewart's (1927, pi. 29, fig. 3) art (1927) synonomized Spirocrypta with Crepidula, show the shelf. Gabb's figure is a fascimile and Stewart's Gabb's description of Spirocrypta recognizes this very is a photograph that has been reproduced in other dis- important characteristic, which helps to distinguish Spi- cussions of Spirocrypta (e.g., Wenz, 1940: 903, fig. rocrypta from Crepidula. In modern Crepidula forni- 2660a). Gabb's and Stewart's figures are based on lecto- cata, the shelf does not spiral into the whorl apex. type ANSP 4221, but, unfortunately, the shelf of this Aperture/shelf features of Spirocrypta pileum and S. specimen is broken. Both figures create the false impres- inomata resemble those of the early Paleocene Spirogal- sion that there is a sinus near both ends of the shelf and erus lamellaria Finlay and Marwick, 1937, from New that the middle part protrudes and is concave. -

Biogeography and Genetic Diversity of the Atlantid Heteropods T ⁎ Deborah Wall-Palmera,B, , Alice K
Progress in Oceanography 160 (2018) 1–25 Contents lists available at ScienceDirect Progress in Oceanography journal homepage: www.elsevier.com/locate/pocean SCCWRP #1038 Biogeography and genetic diversity of the atlantid heteropods T ⁎ Deborah Wall-Palmera,b, , Alice K. Burridgeb,c, Erica Goetzed, Frank R. Stokvisb, Arie W. Janssenb, Lisette Mekkesb,c, María Moreno-Alcántarae, Nina Bednaršekf, Tom Schiøtteg, Martin Vinther Sørenseng, Christopher W. Smarta, Katja T.C.A. Peijnenburgb,c a School of Geography, Earth and Environmental Sciences, University of Plymouth, Plymouth PL4 8AA, UK b Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands c Institute for Biodiversity and Ecosystem Dynamics (IBED), University of Amsterdam, 1090 GE Amsterdam, The Netherlands d Department of Oceanography, University of Hawai‘iatMānoa, Honolulu, HI 96822, USA e Departamento de Plancton y Ecología Marina, Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz C.P. 23096, Mexico f Southern California Coastal Waters Research Project, Harbor Blvd #110, Costa Mesa, CA 92626, USA g The Natural History Museum of Denmark, University of Copenhagen, 2100 Copenhagen, Denmark ARTICLE INFO ABSTRACT Keywords: The atlantid heteropods are regularly encountered, but rarely studied marine planktonic gastropods. Relying Atlantidae on a small (< 14 mm), delicate aragonite shell and living in the upper ocean means that, in common with Atlanta pteropods, atlantids are likely to be affected by imminent ocean changes. Variable shell morphology and Cytochrome c oxidase subunit 1 (mtCO1) widespread distributions indicate that the family is more diverse than the 23 currently known species. DNA barcoding Uncovering this diversity is fundamental to determining the distribution of atlantids and to understanding Planktonic gastropods their environmental tolerances.