BIOLOGICALLY ACTIVE COMPOUNDS of KNOTWEED (Reynoutria Spp
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Japanese Knotweed Fallopia Japonica (Houtt.) R. Decr. Or Polygonum Cuspidatum Sieb
Japanese knotweed Fallopia japonica (Houtt.) R. Decr. or Polygonum cuspidatum Sieb. & Zucc. Giant knotweed Fallopia sachalinensis (F. Schmidt ex Maxim.) R. Decr. or Polygonum sachalinense F. Schmidt ex Maxim. Bohemian knotweed Fallopia × bohemica (Chrtek & Chrtková) J. P. Bailey or Polygonum ×bohemicum (J. Chrtek & Chrtkovß) Zika & Jacobson [cuspidatum ×sachalinense] Family: Polygonaceae Synonyms for Fallopia japonica: Pleuropterus cuspidatus (Sieb. & Zucc.) Moldenke, P. zuccarinii (Small) Small, Polygonum cuspidatum Sieb. & Zucc. var. compactum (Hook. f.) Bailey, P. zuccarinii Small, Reynoutria japonica Houtt. Other common names: Japanese bamboo, fleeceflower, Mexican bamboo Synonyms for Fallopia sachalinensis: Reynoutria sachalinensis (F. Schmidt ex Maxim.) Nakai, Tiniaria sachalinensis (F. Schmidt) Janchen Other common names: none Synonyms for Fallopia x bohemica: none Other common names: none Invasiveness Rank: 87 The invasiveness rank is calculated based on a species’ ecological impacts, biological attributes, distribution, and response to control measures. The ranks are scaled from 0 to 100, with 0 representing a plant that poses no threat to native ecosystems and 100 representing a plant that poses a major threat to native ecosystems. Description Japanese knotweed is a perennial plant that grows from long, creeping rhizomes. Rhizomes are thick, extensive, and 5 to 6 meters long. They store large quantities of carbohydrates. Stems are stout, hollow reddish-brown, swollen at the nodes, and 1 ¼ to 2 ¾ meters tall. Twigs often zigzag slightly from node to node. Leaves are alternate, 5 to 15 cm long, and broadly ovate with more or less truncate bases and acuminate tips. They have short petioles. Plants are dioecious, with male and female flowers on separate plants. Inflorescences are many-flowered, branched, open, and lax. -

Stace Edition 4: Changes
STACE EDITION 4: CHANGES NOTES Changes to the textual content of keys and species accounts are not covered. "Mention" implies that the taxon is or was given summary treatment at the head of a family, family division or genus (just after the key if there is one). "Reference" implies that the taxon is or was given summary treatment inline in the accounts for a genus. "Account" implies that the taxon is or was given a numbered account inline in the numbered treatments within a genus. "Key" means key at species / infraspecific level unless otherwise qualified. "Added" against an account, mention or reference implies that no treatment was given in Edition 3. "Given" against an account, mention or reference implies that this replaces a less full or prominent treatment in Stace 3. “Reduced to” against an account or reference implies that this replaces a fuller or more prominent treatment in Stace 3. GENERAL Family order changed in the Malpighiales Family order changed in the Cornales Order Boraginales introduced, with families Hydrophyllaceae and Boraginaceae Family order changed in the Lamiales BY FAMILY 1 LYCOPODIACEAE 4 DIPHASIASTRUM Key added. D. complanatum => D. x issleri D. tristachyum keyed and account added. 5 EQUISETACEAE 1 EQUISETUM Key expanded. E. x meridionale added to key and given account. 7 HYMENOPHYLLACEAE 1 HYMENOPHYLLUM H. x scopulorum given reference. 11 DENNSTAEDTIACEAE 2 HYPOLEPIS added. Genus account added. Issue 7: 26 December 2019 Page 1 of 35 Stace edition 4 changes H. ambigua: account added. 13 CYSTOPTERIDACEAE Takes on Gymnocarpium, Cystopteris from Woodsiaceae. 2 CYSTOPTERIS C. fragilis ssp. fragilis: account added. -

Separation of Hydroxyanthraquinones by Chromatography
Separation of hydroxyanthraquinones by chromatography B. RITTICH and M. ŠIMEK* Research Institute of Animal Nutrition, 691 23 Pohořelice Received 6 May 1975 Accepted for publication 25 August 1975 Chromatographic properties of hydroxyanthraquinones have been examined. Good separation was achieved using new solvent systems for paper and thin-layer chromatography on common and impregnated chromatographic support materials. Commercial reagents were analyzed by the newly-developed procedures. Было изучено хроматографическое поведение гидроксиантрахинонов. Хорошее разделение было достигнуто при использовании предложенных новых хроматографических систем: бумажная хроматография смесью уксусной кислоты и воды на простой бумаге или бумаге импрегнированной оливковым мас лом, тонкослойная хроматография на целлюлозе импрегнированной диметилфор- мамидом и на силикагеле без или с импрегнацией щавелевой или борной кислотами. Anthraquinones constitute an important class of organic substances. They are produced industrially as dyes [1] and occur also in natural products [2]. The fact that some hydroxyanthraquinones react with metal cations to give colour chelates has been utilized in analytical chemistry [3]. Anthraquinone and its derivatives can be determined spectrophotometrically [4—6] and by polarography [4]. The determination of anthraquinones is frequently preceded by a chromatographic separation the purpose of which is to prepare a chemically pure substance. For chromatographic separation of anthraquinone derivatives common paper [7—9] and paper impregnated with dimethylformamide or 1-bromonaphthalene has been used [10, 11]. Dyes derived from anthraquinone have also been chromatographed on thin layers of cellulose containing 10% of acetylcellulose [12]. Thin-layer chromatography on silica gel has been applied in the separation of dihydroxyanthraquinones [13], di- and trihydroxycarbox- ylic acids of anthraquinones [14] and anthraquinones occurring in nature [15]. -

Asclepias Syriaca L.) After a Single Herbicide Treatment in Natural Open Sand Grasslands László Bakacsy* & István Bagi
www.nature.com/scientificreports OPEN Survival and regeneration ability of clonal common milkweed (Asclepias syriaca L.) after a single herbicide treatment in natural open sand grasslands László Bakacsy* & István Bagi Invasive species are a major threat to biodiversity, human health, and economies worldwide. Clonal growth is a common ability of most invasive plants. The clonal common milkweed Asclepias syriaca L. is the most widespread invasive species in Pannonic sand grasslands. Despite of being an invader in disturbed semi-natural vegetation, this plant prefers agricultural felds or plantations. Herbicide treatment could be one of the most cost-efective and efcient methods for controlling the extended stands of milkweed in both agricultural and protected areas. The invasion of milkweed stand was monitored from 2011 to 2017 in a strictly protected UNESCO biosphere reserve in Hungary, and a single herbicide treatment was applied in May 2014. This single treatment was successful only in a short-term but not in a long-term period, as the number of milkweed shoots decreased following herbicide treatment. The herbicide translocation by rhizomatic roots induced the damage of dormant bud banks. The surviving buds developing shoots, growth of the milkweed stand showed a slow regeneration for a longer-term period. We concluded that the successful control of milkweed after herbicide treatment depends on repeated management of treated areas to suppress further spreading during subsequent seasons. Currently, invasive species are a major threat to biodiversity, human health, and economies 1–4. It has been esti- mated that the fght against invasive species and the damage caused by them in European Union accounts for a minimum of 9.6–12.7 billion euros annually, and this amount is expected to rise to 20 billion euros annually1,5–7. -

UHPLC Analysis of Reynoutria Japonica Houtt. Rhizome Preparations Regarding Stilbene and Anthranoid Composition and Their Antimycobacterial Activity Evaluation
plants Article UHPLC Analysis of Reynoutria japonica Houtt. Rhizome Preparations Regarding Stilbene and Anthranoid Composition and Their Antimycobacterial Activity Evaluation Fabian Alperth, Lena Melinz, Johannes-Paul Fladerer and Franz Bucar * Institute of Pharmaceutical Sciences, University of Graz, Beethovenstraße 8, 8010 Graz, Austria; [email protected] (F.A.); [email protected] (L.M.); johannes.fl[email protected] (J.-P.F.) * Correspondence: [email protected]; Tel.: +43-316-380-5531 Abstract: Reynoutria japonica Houtt. is a critical invasive alien plant in Europe and North America with a drastic impact on native flora. However, R. japonica has medicinal potential, especially as a source of stilbenes. In order to explore the potential of simple extractions of R. japonica, we conducted qualitative and quantitative analyses of fresh R. japonica rhizome infusion, decoction, and macerates with ethanol by UHPLC-DAD-ESI-MSn and UHPLC-DAD, with a focus on major constituent groups of stilbenes and anthranoids. Since R. japonica rhizome extracts showed antimicrobial potential in the past, we also evaluated the antimycobacterial effect of raw R. japonica extracts for the first time against Mycobacterium smegmatis. Of thirty-four characterized substances, six were stilbenes and twelve anthranoids. The main constituents, four trans-stilbenes and eight anthranoids, were quantified in a validated UHPLC-DAD method. The 38% ethanol macerate showed high stilbene (155.078 mg/100 g Citation: Alperth, F.; Melinz, L.; fluid extract) and low anthranoid content (5.420 mg/100 g fluid extract), while decoction showed the Fladerer, J.-P.; Bucar, F. UHPLC µ Analysis of Reynoutria japonica Houtt. highest anthranoids. -

Reynoutria Japonica Houtt. © Morvant Y
Reynoutria japonica Houtt. © Morvant Y. CBN Méditerranéen de Porquerolles La Renouée du Japon Plantae, Spermatophytes, Angiospermes, Dicotylédones, Caryophyllales, Polygonaceae Synonymes : Fallopia japonica (Houtt.) Ronse Decraene Polygonum cuspidatum Siebold & Zucc. Polygonum reynoutria Makino Polygonum sieboldii Reinw. ex de Vries Polygonum zuccarinii Small Pleuropterus cuspidatus (Siebold & Zucc.) H. Gross Tiniaria cuspidata (Houtt.) Hedberg Fiche réalisée par la Fédération des Conservatoires botaniques nationaux Description générale Plante herbacée, vivace rhizomateuse, à port buissonnant. Annuellement, elle forme des tiges aériennes robustes, souvent tachetées de rouge (forme de lenticelle) et pouvant atteindre 3 m de hauteur et 4 cm de diamètre, formant de vastes massifs denses. Elle possède des rhizomes, tiges souterraines bien développées et lignifiées, assurant la pérennité de la plante et permettant la reproduction végétative. Elles peuvent atteindre 15-20 m de long et pénétrer dans le sol jusqu'à 2-3 m de profondeur. Des racines adventives sont émises des rhizomes. Son limbe foliaire est largement ovale, atteignant 20 cm de long et est brusquement tronqué à la base. Les fleurs de couleur blanc-crème et blanc verdâtre se développent en panicule plus ou moins lâches de 8-12 cm de longueur. Biologie/Ecologie Reproduction Plante dioïque (pied mâle et pied femelle) à floraison automnale (août-octobre) et pollinisation entomophile. Reproduction sexuée : En France, seuls des individus mâles stériles sont connus sur le territoire. La formation de graines est réalisée par fécondation croisée avec le pollen de Reynoutria sachalinensis, donnant naissance à un hybride Reynoutria x bohemica. La production de graines viables est rare et les plantules sont généralement bloquées dans leur développement. -
Vegetation and Ecological Characterisitics of Mixed-Conifer
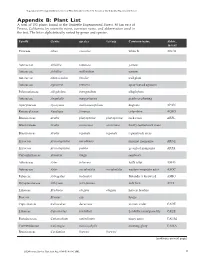
Vegetation and Ecological Charactistics of Mixed-Conifer and Red Fir Forests at the Teakettle Experimental Forest Vegetation and Ecological Characteristics of Mixed-Conifer and Red Fir Forests at the Teakettle Experimental Forest Appendix B: Plant List A total of 152 plants found at the Teakettle Experimental Forest, 80 km east of Fresno, California, by scientific name, common name, and abbreviation used in the text. The list is alphabetically sorted by genus and species. Family Genus species var/ssp Common name Abbre. in text Pinaceae Abies concolor white fir ABCO Pinaceae Abies magnifica red fir ABMA Asteraceae Achillea lanulosa yarrow Asteraceae Achillea millefolium yarrow Asteraceae Adenocaulon bicolor trail plant Asteraceae Agroseris retrorsa spear-leaved agoseris Polemoniaceae Allophylum intregifolium allophylum Asteraceae Anaphalis margaritacea pearly everlasting Apocynaceae Apocynum androsaemifolium dogbane APAN Ranunculaceae Aquilegia formosa columbine AQFO Brassicaceae Arabis platysperma platysperma rock cress ARPL Brassicaceae Arabis rectissima rectissima bristly-leaved rock cress Brassicaceae Arabis repanda repanda repand rock cress Ericaceae Arctostaphylus nevadensis pinemat manzanita ARNE Ericaceae Arctostaphylus patula greenleaf manzanita ARPA Caryophyliaceae Arenaria kingii sandwort Asteraceae Aster foliaceus leafy aster ASFO Asteraceae Aster occidentalis occidentalis western mountain aster ASOC Fabaceae Astragalus bolanderi Bolander’s locoweed ASBO Dryopteridaceae Athryium felix-femina lady fern ATFI Liliaceae Brodiaea elegans elegans harvest brodeia Poaceae Bromus ssp. brome Cupressaceae Calocedrus decurrens incense cedar CADE Liliaceae Calochortus leichtlinii Leichtlin’s mariposa lily CALE Portulacaceae Calyptridium umbellatum pussy paws CAUM Convuvulaceae Calystegia malacophylla morning glory CAMA Brassicaceae Cardamine breweri breweri (continues on next page) 46 USDA Forest Service Gen.Tech. Rep. PSW-GTR-186. 2002. USDA Forest Service Gen.Tech. Rep. -

Natural Hydroxyanthraquinoid Pigments As Potent Food Grade Colorants: an Overview
Review Nat. Prod. Bioprospect. 2012, 2, 174–193 DOI 10.1007/s13659-012-0086-0 Natural hydroxyanthraquinoid pigments as potent food grade colorants: an overview a,b, a,b a,b b,c b,c Yanis CARO, * Linda ANAMALE, Mireille FOUILLAUD, Philippe LAURENT, Thomas PETIT, and a,b Laurent DUFOSSE aDépartement Agroalimentaire, ESIROI, Université de La Réunion, Sainte-Clotilde, Ile de la Réunion, France b LCSNSA, Faculté des Sciences et des Technologies, Université de La Réunion, Sainte-Clotilde, Ile de la Réunion, France c Département Génie Biologique, IUT, Université de La Réunion, Saint-Pierre, Ile de la Réunion, France Received 24 October 2012; Accepted 12 November 2012 © The Author(s) 2012. This article is published with open access at Springerlink.com Abstract: Natural pigments and colorants are widely used in the world in many industries such as textile dying, food processing or cosmetic manufacturing. Among the natural products of interest are various compounds belonging to carotenoids, anthocyanins, chlorophylls, melanins, betalains… The review emphasizes pigments with anthraquinoid skeleton and gives an overview on hydroxyanthraquinoids described in Nature, the first one ever published. Trends in consumption, production and regulation of natural food grade colorants are given, in the current global market. The second part focuses on the description of the chemical structures of the main anthraquinoid colouring compounds, their properties and their biosynthetic pathways. Main natural sources of such pigments are summarized, followed by discussion about toxicity and carcinogenicity observed in some cases. As a conclusion, current industrial applications of natural hydroxyanthraquinoids are described with two examples, carminic acid from an insect and Arpink red™ from a filamentous fungus. -

Characteristics That Make the Fallopia Genus (Polygonaceae) Highly Invasive
Ecological Questions 16/2012: 23 – 27 DOI: 10.2478/v10090-012-0002-6 Characteristics that make the Fallopia genus (Polygonaceae) highly invasive Justyna Sołtysiak, Teresa Brej Department of Botany and Plant Ecology, Wrocław University of Environmental and Life Sciences, pl. Grunwaldzki 24a, 50–363 Wrocław, Poland e-mail: [email protected] Abstract. Representatives of the Fallopia genus: Fallopia japonica, Fallopia sachalinensis and Fallopia × bohemica are known as successful invaders, wide spread throughout Europe and North America. This paper focuses on the invasive Fallopia complex and presents some features (a wide ecological amplitude, high competition abilities, sexual reproduction by hybridization) responsible for the fact that all species of the Fallopia genus are aggressive and noxious invaders. Key words: plant invasion, Fallopia japonica, Fallopia sachalinensis, Fallopia × bohemica. 1. Introduction cies of exotic plants have been introduced as a crop and have escaped to become established in natural ecosystems Biological invasions belong to the main problems of con- (Pimentel et al. 2007). temporary ecology and they are considered as a signifi- Recent research were dedicate to invasive species, de- cant component of global changes, connected with human spite that fact scientist still try to find the answer for the activity (Vitousek et al. 1997; Vilá et al. 2007; McKinney question: what characteristics make them invasive? (Rej- 2006). mánek 1995). The major point of the biological invasions was the The article is dedicated to the Fallopia genus, whose discovery of America by Christopher Columbus in 1492, some species are known as successful invaders, wide which has facilitated exchange of goods between new and spread in Europe and North America. -

Fallopia Japonica – Japanese Knotweed
Fallopia japonica – Japanese knotweed Japanese knotweed, sometimes referred to What is it? as donkey rhubarb for its sour red spring shoots, is a perennial plant in the Buckwheat family (Polygonaceae). It has large broad green leaves; tall, thick, sectioned and somewhat reddish zigzagging stems; and racemes of small papery flowers in summer. Photo by Liz West 2007 Other scientific names (synonyms) for Japanese knotweed are Reynoutria japonica and Polygonum cuspidatum. When does it grow? Shoots emerge from rhizomes (modified underground stems) from late March to mid-April. A spring freeze or deep frost can top kill new growth, but new shoots readily crop up from the hardy rootstalks. Growth continues rapidly once the weather begins to warm reaching heights up to 10 feet or greater by summer. R. Buczynski 2020 4.15.2020 Where is it from? Japanese knotweed is native to eastern Asia and was introduced to the United Kingdom in the 1800’s as a vigorous garden ornamental. Before becoming illegal to plant in England it was horticulturally introduced from the UK to the United States. Where is it now? Japanese knotweed has been reported extensively in the Northeastern U. S. and is currently present in all three counties (Hunterdon, Morris, and Somerset) within the upper Raritan watershed where it continues to spread into moist disturbed areas along waterways. Photo by Roger Kidd © Why is it invasive? Although knotweed can spread by seed, it is most effective at spreading underground via rhizomes that extend outward as well as downward, producing new shoots up to 70 feet away. If detached from the plant, small fragments of rhizome can survive and produce new plants wherever they land. -

Pocket Guide for Western North Carolina Partnership (SACWMP), 2011
DO NOT BUY Invasive Exotic Plant List Produced by the Southern Appalachian Cooperative Weed Management pocket guide for western north carolina Partnership (SACWMP), 2011 Western North Carolina has to offer! offer! to has Carolina North Western ) allegheniensis Rubus do not buy these invasives buy natives or alternatives ( Blackberry Allegheny ) alba Quercus ( Oak White of beautiful native plants that that plants native beautiful of ! Mimosa (Silk Tree) Albizia julibrissin Common Serviceberry (Amelanchier arborea) ) nigra Juglans ( Walnut Black Eastern Eastern Redbud (Cercis canadensis) multitude the enjoy and environment, To use your pocket guide: ) virginiana Diospyros ( Persimmon Flowering Dogwood (Cornus florida) whole the of quality the to Add counts. ) pumila Castanea ( Chinquapin the environment a favor on both both on favor a environment the 1 Print on letter-size paper. Japanese Barberry Berberis thunbergii Mountain Pepperbush (Clethra acuminata) wildlife for great Virginia Sweetspire (Itea virginica) doing are you plants, native planting Spicebush (Lindera benzoin) By habitat. species’ of loss the and 2 Cut along outer black line. are the spread of invasive exotic plants plants exotic invasive of spread the are ) fistulosum Eupatorium Butterfly Bush Buddleia davidii Swamp Milkweed (Asclepias incarnata) ( Weed Pye Joe ) ) purpurea (Echinacea Coneflower Purple Purple Coneflower (Echinacea purpurea) Carolina North Western in problems 3 Fold on dotted blue lines. ) syriaca Asclepias ( Milkweed Common Joe Pye Weed (Eupatorium fistulosum) -

United States Patent (19) 11 Patent Number: 4,746,461 Zielske (45) Date of Patent: May 24, 1988
United States Patent (19) 11 Patent Number: 4,746,461 Zielske (45) Date of Patent: May 24, 1988 54 METHOD FOR PREPARING 4,041,051 8/1977 Yamado et al. ..................... 260/371 1,4-DIAMINOANTHRAQUINONES AND 4,661,293 4/1987 Zielske ................................ 260/377 INTERMEDIATES THEREOF FOREIGN PATENT DOCUMENTS 75 Inventor: Alfred G. Zielske, Pleasanton, Calif. 2014178 8/1979 United Kingdom................ 260/377 2019870 1 1/1979 United Kingdom ..... ... 260/377 73 Assignee: The Clorox Company, Oakland, Calif. 2100307A 5/1980 United Kingdom ................ 260/377 (21) Appl. No.: 945,906 OTHER PUBLICATIONS 22 Filed: Dec. 23, 1986 Morrison & Boyd, Organic Chemistry, 3rd ed., 1973, pp. 458, 456,527, 528. s Related U.S. Application Data Chemical Abstract, vol. 88, #10498g&h, Schultz et al., 60 Division of Ser. No. 868,884, May 23, 1986, Pat. No. 1978, "Synthesis of Meso-Substituted Hydroxyantha 4,661,293, which is a continuation of Ser. No. 556,835, rones with Laxative Activity Parts I & II' Arch. Dec. 1, 1983, abandoned. Pharm, vol. 310, No. 10, pp. 769-780, 1977. 51 Int. Cl." ...................... C11D 15/02; C11D 13/00 Primary Examiner-Glennon H. Hollrah 52 U.S. C. .................................... 260/370; 260/371; Assistant Examiner-Raymond Covington 260/374 Attorney, Agent, or Firm-Majestic, Gallagher, Parsons 58 Field of Search ............... 260/370, 371, 377, 378, & Siebert 260/374 (57) ABSTRACT (56) References Cited Aminoanthraquinones are generally useful as dyes and U.S. PATENT DOCUMENTS coloring agents. But known methods of synthesizing 2,174,751 10/1939 Koerberle ........................... 260/367 unsymmetrically substituted 1,4-diaminoanthraquinones 2,183,652 12/1939 Lord ............