Complete Issue
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Net Zero by 2050 a Roadmap for the Global Energy Sector Net Zero by 2050
Net Zero by 2050 A Roadmap for the Global Energy Sector Net Zero by 2050 A Roadmap for the Global Energy Sector Net Zero by 2050 Interactive iea.li/nzeroadmap Net Zero by 2050 Data iea.li/nzedata INTERNATIONAL ENERGY AGENCY The IEA examines the IEA member IEA association full spectrum countries: countries: of energy issues including oil, gas and Australia Brazil coal supply and Austria China demand, renewable Belgium India energy technologies, Canada Indonesia electricity markets, Czech Republic Morocco energy efficiency, Denmark Singapore access to energy, Estonia South Africa demand side Finland Thailand management and France much more. Through Germany its work, the IEA Greece advocates policies Hungary that will enhance the Ireland reliability, affordability Italy and sustainability of Japan energy in its Korea 30 member Luxembourg countries, Mexico 8 association Netherlands countries and New Zealand beyond. Norway Poland Portugal Slovak Republic Spain Sweden Please note that this publication is subject to Switzerland specific restrictions that limit Turkey its use and distribution. The United Kingdom terms and conditions are available online at United States www.iea.org/t&c/ This publication and any The European map included herein are without prejudice to the Commission also status of or sovereignty over participates in the any territory, to the work of the IEA delimitation of international frontiers and boundaries and to the name of any territory, city or area. Source: IEA. All rights reserved. International Energy Agency Website: www.iea.org Foreword We are approaching a decisive moment for international efforts to tackle the climate crisis – a great challenge of our times. -

Caving: Safety Activity Checkpoints
Caving: Safety Activity Checkpoints Caving—also called “spelunking” (speh-LUNK-ing) is an exciting, hands-on way to learn about speleology (spee-lee-AH- luh-gee), the study of caves, in addition to paleontology (pay-lee-en-TAH-luh-gee), the study of life from past geologic periods by examining plant and animal fossils. As a sport, caving is similar to rock climbing, and often involves using ropes to crawl and climb through cavern nooks and crannies. These checkpoints do not apply to groups taking trips to tourist or commercial caves, which often include safety features such as paths, electric lights, and stairways. Girl Scout Daisies and Brownies do not participate in caving. Know where to go caving. Connect with your Girl Scout council for site suggestions. Also, the National Speleological Society provides an online search tool for U.S. caving clubs, and the National Park Service provides information about National Park caves. Include girls with disabilities. Communicate with girls with disabilities and/or their caregivers to assess any needs and accommodations. Learn more about the resources and information that the National Center on Accessibility and the National Center of Physical Activities and Disabilities provide to people with disabilities. Caving Gear Basic Gear Sturdy boots with ankle protection (hiking boots for dry areas; rubber boots or wellies for wet caves) Warm, rubber gloves (to keep hands warm and protect against cuts and abrasions) Nonperishable, high-energy foods such as fruits and nuts Water Specialized Gear -

Lehman Caves Management Plan
National Park Service U.S. Department of the Interior Great Basin National Park Lehman Caves Management Plan June 2019 ON THE COVER Photograph of visitors on tour of Lehman Caves NPS Photo ON THIS PAGE Photograph of cave shields, Grand Palace, Lehman Caves NPS Photo Shields in the Grand Palace, Lehman Caves. Lehman Caves Management Plan Great Basin National Park Baker, Nevada June 2019 Approved by: James Woolsey, Superintendent Date Executive Summary The Lehman Caves Management Plan (LCMP) guides management for Lehman Caves, located within Great Basin National Park (GRBA). The primary goal of the Lehman Caves Management Plan is to manage the cave in a manner that will preserve and protect cave resources and processes while allowing for respectful recreation and scientific use. More specifically, the intent of this plan is to manage Lehman Caves to maintain its geological, scenic, educational, cultural, biological, hydrological, paleontological, and recreational resources in accordance with applicable laws, regulations, and current guidelines such as the Federal Cave Resource Protection Act and National Park Service Management Policies. Section 1.0 provides an introduction and background to the park and pertinent laws and regulations. Section 2.0 goes into detail of the natural and cultural history of Lehman Caves. This history includes how infrastructure was built up in the cave to allow visitors to enter and tour, as well as visitation numbers from the 1920s to present. Section 3.0 states the management direction and objectives for Lehman Caves. Section 4.0 covers how the Management Plan will meet each of the objectives in Section 3.0. -

Junior Cave Scientist Cave and Karst Program Activity Book Ages 5 – 12+
National Park Service U.S. Department of the Interior Geologic Resources Division Junior Cave Scientist Cave and Karst Program Activity Book Ages 5 – 12+ Name: Age: Explore • Learn • Protect 1 Become a Junior Cave Scientist Caves and karst landscapes are found throughout the United States. These features are important as part of our Nation's geologic heritage. In this book, you will explore a fascinating and fragile underground world, learn about the values of caves and karst landscapes, and complete fun educational activities. Explore magnificent and beautiful caves. You will find an amazing underground world just beneath your feet! Learn about caves and karst systems and the work that cave scientists do. Protect our natural environments and the things that make caves and karst areas special. To earn your badge, complete at least activities. (Your Age) Activities in this book are marked with an age indicator. Look for the symbols below: Flashlight Lantern Helmet and Headlamp Ages 5 - 7 Ages 8 – 11 Ages 12 and Older Put a check next to your age indicator on each page that you complete. I received this book from: After completing the activities, there are two ways to receive your Junior Cave Scientist badge: • Return the completed book to a ranger at a participating park, or 2 • Visit go.nps.gov/jrcavesci What are Speleo-Fact: Mammoth Cave is the longest cave in world with over 405 miles (652 km) of connected passageways. Caves and Karst? Caves are naturally occurring voids, cavities, interconnected passageways, or alcoves in the earth. Caves preserve fossils, minerals, ecosystems, and records of past climates. -

Mountains, Streams, and Lakes of Oklahoma I
Information Series #1, June 1998 Mountains, Streams, and Lakes of OklahomaI Kenneth S. Johnson2 INTRODUCTION valleys, hills, and plains throughout most of the re mainder of Oklahoma (Fig. 1). All the major lakes and Mountains and streams define the landscape of reservoirs of Oklahoma are man-made, and they are Oklahoma (Fig. 1). The mountains consist mainly of important for flood contr()l, water supply, recreation, resistant rock masses that were folded, faulted, and and generation of hydroelectric power. Natural lakes thrust upward in the geologic past (Fig. 2), whereas in Oklahoma are limited to oxbow lakes along major the streams have persisted in eroding less-resistant streams and to playa lakes in the High Plains region rock units and lowering the landscape to form broad of the west. Alphabetical List of20 Lakes with Largest Surface Area (from Oklahoma Water Atlas, Oklahoma Water Resources Board) 1. Broken Bow 11. Lake 0' The Cherokees 2. Canton 12. Oologah 3. Eufaula 13. Robert s. Kerr 4. Fort Gibson 14. Sardis 5. Foss 15. Skiatook 6. Great Salt Plains 16. Tenkiller Ferry ·7. Hudson 17. Texoma 8. Hugo 18. Waurika 9. Kaw 19. Webbers Falls 10. Keystone 20. Wister Modified from Historical Atlas of Oklahoma, by John W. Morris, Charles R. Goins, and Edwin C. 25 McReynolds. Copyright © 1986 by the University I of Oklahoma Press. o 40 80Km Figure 1. Mountains, streams, and principal lakes of Oklahoma. lReprinted from Oklahoma Geology Notes (1993), vol. 53, no. 5, p. 180-188. The Notes article was reprinted and expanded from Oklahoma Almanac, 1993-1994, Oklahoma Department of Lihraries, p. -

NSS Conservation and Preservation Policies ( .Pdf )
Part 2-Conservation, Management, Ethics: NSS Conservation and Preservation Policies 253 Section C-Improving Caver Ethics NSS Conservation and Preservation Policies NSS Cave Conservation Policy The National Speleological Society believes: Caves have unique scientific, recreational, and scenic values. These values are endangered by both carelessness and intentional vandalism. These values, once gone, cannot be recovered. The responsibility for protecting caves must be assumed by those who study and enjoy them. Accordingly, the intention of the Society is to work for the preservation of caves with a realistic policy supported by effective programs for: Encouraging self-discipline among cavers. Education and research concerning the causes and prevention of cave damage. Special projects, including cooperation with other groups similarly dedicated to the conservation of natural areas. Specifically: All contents of a cave-formations, life, and loose deposits-are significant for their enjoyment and interpretation. Caving parties should leave a cave as they find it. Cavers should provide means for the removal of waste. Cavers should limit marking to a few, small, removable signs as needed for surveys. Cavers should especially exercise extreme care not to accidentally break or soil formations, disturb life forms, or unnecessarily increase the number of disfiguring paths through an area. Scientific collection is professional, selective, al)d minimal. The collecting of mineral or biological material for display purposes-including previ- ously broken or dead specimens-is never justified, as it encourages others to collect and destroy the interest of the cave. The Society encourages projects such as: Establishing cave preserves. Placing entrance gates where appropriate. Opposing the sale of speleothems. -

5 Years on Ice Age Europe Network Celebrates – Page 5
network of heritage sites Magazine Issue 2 aPriL 2018 neanderthal rock art Latest research from spanish caves – page 6 Underground theatre British cave balances performances with conservation – page 16 Caves with ice age art get UnesCo Label germany’s swabian Jura awarded world heritage status – page 40 5 Years On ice age europe network celebrates – page 5 tewww.ice-age-europe.euLLING the STORY of iCe AGE PeoPLe in eUROPe anD eXPL ORING PLEISTOCene CULtURAL HERITAGE IntrOductIOn network of heritage sites welcome to the second edition of the ice age europe magazine! Ice Age europe Magazine – issue 2/2018 issn 25684353 after the successful launch last year we are happy to present editorial board the new issue, which is again brimming with exciting contri katrin hieke, gerdChristian weniger, nick Powe butions. the magazine showcases the many activities taking Publication editing place in research and conservation, exhibition, education and katrin hieke communication at each of the ice age europe member sites. Layout and design Brightsea Creative, exeter, Uk; in addition, we are pleased to present two special guest Beate tebartz grafik Design, Düsseldorf, germany contributions: the first by Paul Pettitt, University of Durham, cover photo gives a brief overview of a groundbreaking discovery, which fashionable little sapiens © fumane Cave proved in february 2018 that the neanderthals were the first Inside front cover photo cave artists before modern humans. the second by nuria sanz, water bird – hohle fels © urmu, director of UnesCo in Mexico and general coordi nator of the Photo: burkert ideenreich heaDs programme, reports on the new initiative for a serial transnational nomination of neanderthal sites as world heritage, for which this network laid the foundation. -

Geographical Names and Sustainable Tourism
No. 59 NOVEMBERNo. 59 NOVEMBER 2020 2020 Geographical Names and Sustainable Tourism Socio- Institutional cultural Sustainable Tourism Economic Environmental Table of Contents The Information Bulletin of the United Nations MESSAGE FROM THE CHAIRPERSON ............................................... 3 Group of Experts on Geographical Names (formerly Reconsidérer notre mobilité ......................................................... 3 UNGEGN Newsletter) is issued twice a year by the Secretariat of the Group of Experts. The Secretariat Reconsider our mobility ............................................................... 4 is served by the Statistics Division (UNSD), MESSAGE FROM THE SECRETARIAT ................................................. 5 Department for Economic and Social Affairs (DESA), Secretariat of the United Nations. Contributions “Geographical names and sustainable tourism ............................ 5 and reports received from the Experts of the Group, IN MEMORIAM ................................................................................ 7 its Linguistic/Geographical Divisions and its Working Groups are reviewed and edited jointly by the Danutė Janė Mardosienė (1947-2020) ........................................ 7 Secretariat and the UNGEGN Working Group on SPECIAL FEATURE: GEOGRAPHICAL NAMES AND SUSTAINABLE Publicity and Funding. Contributions for the TOURISM ......................................................................................... 9 Information Bulletin can only be considered when they are made -

Cave Research Foundation (CRF) Is a 501(C)(3) Organization Founded in Kentucky in 1957 for the Purpose of Facilitating Cave Research
CAVE RESEARCH FOUNDATION Ozarks Operation Area by Scott House The Cave Research Foundation (CRF) is a 501(c)(3) organization founded in Kentucky in 1957 for the purpose of facilitating cave research. Today it continues its mission with operations across the country and in expeditions around the world. The CRF is administered by officers, a board and an operations council. An operations manager is appointed by the CRF board to coordinate and oversee activities within an operation. Other individuals within the operation may head specific projects or act as functionaries of one sort or another. Following is a brief description of various CRF Ozarks Operation projects. Mark Twain National Forest: CRF works with Mark Twain National Forest (USFS) through a series of cooperative agreements and modifications. These cooperative agreements superseded challenge cost-share agreements and volunteer agreements. Cave Mapping and Biological Inventory: This has been our largest and longest running project. It originated in 1990 with a management concern centered on an application by the Doe Run Company for mineral prospecting within the Eleven Point and Current River watersheds. The aim was to document all caves within the 250 square kilometer mineral lease area by detailed mapping, biological inventory and comprehensive descriptions. More than 120 caves were visited and documented over the 6 year course of the study. Following on from that initial phase, MTNF has continued to provide funding, through one vehicle or another, for an expanded project which is gradually documenting caves throughout the Mark Twain National Forest. Approximately 450 caves have been documented during this ongoing study. -

Mites (Acari, Mesostigmata) from Rock Cracks and Crevices in Rock Labirynths in the Stołowe Mountains National Park (SW Poland)
BIOLOGICAL LETT. 2014, 51(1): 55–62 Available online at: http:/www.degruyter.com/view/j/biolet DOI: 10.1515/biolet-2015-0006 Mites (Acari, Mesostigmata) from rock cracks and crevices in rock labirynths in the Stołowe Mountains National Park (SW Poland) JACEK KAMCZYC and MACIEJ SKORUPSKI Department of Game Management and Forest Protection, Poznań University of Life Sciences, Wojska Polskiego 71C, 60-625 Poznań Corresponding author: Jacek Kamczyc, [email protected] (Received on 7 January 2013; Accepted on 7 April 2014) Abstract: The aim of this study was to recognize the species composition of soil mites of the order Mesostigmata in the soil/litter collected from rock cracks and crevices in Szczeliniec Wielki and Błędne Skały rock labirynths in the area of the Stołowe Mountains National Park (part of the Sudetes in SW Po- land). Overall, 27 species were identified from 41 samples collected between September 2001 and August 2002. The most numerous species in this study were Veigaia nemorensis, Leptogamasus cristulifer, and Gamasellus montanus. Our study has also confirmed the occurrence or rare mite species, such asVeigaia mollis and Paragamasus insertus. Additionally, 5 mite species were recorded as new to the fauna of this Park: Vulgarogamasus remberti, Macrocheles tardus, Pachylaelaps vexillifer, Iphidosoma physogastris, and Dendrolaelaps (Punctodendrolaelaps) eichhorni. Keywords: mesofauna, mites, Mesostigmata, soil, rock cracks, crevices INTRODUCTION The Stołowe Mountains National Park (also known as the Góry Stołowe NP) was established in 1993, in the area of the only table hills in Poland, mainly due to the occurrence of the very specific sandstone landscapes, including rocks labyrinths. The rock labyrinths are generally composed of sandstones blocks, separated by cracks and crevices (Szopka 2002). -

Dear Secretary Salazar: I Strongly
Dear Secretary Salazar: I strongly oppose the Bush administration's illegal and illogical regulations under Section 4(d) and Section 7 of the Endangered Species Act, which reduce protections to polar bears and create an exemption for greenhouse gas emissions. I request that you revoke these regulations immediately, within the 60-day window provided by Congress for their removal. The Endangered Species Act has a proven track record of success at reducing all threats to species, and it makes absolutely no sense, scientifically or legally, to exempt greenhouse gas emissions -- the number-one threat to the polar bear -- from this successful system. I urge you to take this critically important step in restoring scientific integrity at the Department of Interior by rescinding both of Bush's illegal regulations reducing protections to polar bears. Sarah Bergman, Tucson, AZ James Shannon, Fairfield Bay, AR Keri Dixon, Tucson, AZ Ben Blanding, Lynnwood, WA Bill Haskins, Sacramento, CA Sher Surratt, Middleburg Hts, OH Kassie Siegel, Joshua Tree, CA Sigrid Schraube, Schoeneck Susan Arnot, San Francisco, CA Stephanie Mitchell, Los Angeles, CA Sarah Taylor, NY, NY Simona Bixler, Apo Ae, AE Stephan Flint, Moscow, ID Steve Fardys, Los Angeles, CA Shelbi Kepler, Temecula, CA Kim Crawford, NJ Mary Trujillo, Alhambra, CA Diane Jarosy, Letchworth Garden City,Herts Shari Carpenter, Fallbrook, CA Sheila Kilpatrick, Virginia Beach, VA Kierã¡N Suckling, Tucson, AZ Steve Atkins, Bath Sharon Fleisher, Huntington Station, NY Hans Morgenstern, Miami, FL Shawn Alma, -
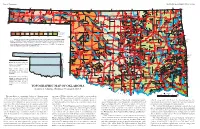
TOPOGRAPHIC MAP of OKLAHOMA Kenneth S
Page 2, Topographic EDUCATIONAL PUBLICATION 9: 2008 Contour lines (in feet) are generalized from U.S. Geological Survey topographic maps (scale, 1:250,000). Principal meridians and base lines (dotted black lines) are references for subdividing land into sections, townships, and ranges. Spot elevations ( feet) are given for select geographic features from detailed topographic maps (scale, 1:24,000). The geographic center of Oklahoma is just north of Oklahoma City. Dimensions of Oklahoma Distances: shown in miles (and kilometers), calculated by Myers and Vosburg (1964). Area: 69,919 square miles (181,090 square kilometers), or 44,748,000 acres (18,109,000 hectares). Geographic Center of Okla- homa: the point, just north of Oklahoma City, where you could “balance” the State, if it were completely flat (see topographic map). TOPOGRAPHIC MAP OF OKLAHOMA Kenneth S. Johnson, Oklahoma Geological Survey This map shows the topographic features of Oklahoma using tain ranges (Wichita, Arbuckle, and Ouachita) occur in southern contour lines, or lines of equal elevation above sea level. The high- Oklahoma, although mountainous and hilly areas exist in other parts est elevation (4,973 ft) in Oklahoma is on Black Mesa, in the north- of the State. The map on page 8 shows the geomorphic provinces The Ouachita (pronounced “Wa-she-tah”) Mountains in south- 2,568 ft, rising about 2,000 ft above the surrounding plains. The west corner of the Panhandle; the lowest elevation (287 ft) is where of Oklahoma and describes many of the geographic features men- eastern Oklahoma and western Arkansas is a curved belt of forested largest mountainous area in the region is the Sans Bois Mountains, Little River flows into Arkansas, near the southeast corner of the tioned below.