Small Rodents and a Lagomorph from the Early Miocene Bukwa Locality, Eastern Uganda
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

The World at the Time of Messel: Conference Volume
T. Lehmann & S.F.K. Schaal (eds) The World at the Time of Messel - Conference Volume Time at the The World The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates 22nd International Senckenberg Conference 2011 Frankfurt am Main, 15th - 19th November 2011 ISBN 978-3-929907-86-5 Conference Volume SENCKENBERG Gesellschaft für Naturforschung THOMAS LEHMANN & STEPHAN F.K. SCHAAL (eds) The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference Frankfurt am Main, 15th – 19th November 2011 Conference Volume Senckenberg Gesellschaft für Naturforschung IMPRINT The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference 15th – 19th November 2011, Frankfurt am Main, Germany Conference Volume Publisher PROF. DR. DR. H.C. VOLKER MOSBRUGGER Senckenberg Gesellschaft für Naturforschung Senckenberganlage 25, 60325 Frankfurt am Main, Germany Editors DR. THOMAS LEHMANN & DR. STEPHAN F.K. SCHAAL Senckenberg Research Institute and Natural History Museum Frankfurt Senckenberganlage 25, 60325 Frankfurt am Main, Germany [email protected]; [email protected] Language editors JOSEPH E.B. HOGAN & DR. KRISTER T. SMITH Layout JULIANE EBERHARDT & ANIKA VOGEL Cover Illustration EVELINE JUNQUEIRA Print Rhein-Main-Geschäftsdrucke, Hofheim-Wallau, Germany Citation LEHMANN, T. & SCHAAL, S.F.K. (eds) (2011). The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates. 22nd International Senckenberg Conference. 15th – 19th November 2011, Frankfurt am Main. Conference Volume. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main. pp. 203. -

Liste Fascicules
LISTE DES FASCICULES PARUS VOLUME 1 Fasc. 1 (1967) J.A. REMY.— Les Palaeotheridae (Perissodactyla) de la faune de mammifères de Fons 1 (Eocène supérieur), 1-46, 20 fig., 12 tabl., 8 pl. Fasc. 2 (1967) J.-L. HARTENBERGER.— Contribution à l'étude de l'anatomie crânienne des rongeurs. I. - Principaux types de cricétodontinés, 47-64, 2 fig., 4 pl. Fasc. 3 (1968) B. SIGÉ.— Les chiroptères du Miocène inférieur de Bouzigues. I. - Etude systématique, 65-133, 28 fig., 10 tabl. Fasc. 4 (1968) J. MICHAUX.— Les Paramyidae (Rodentia) de l'Eocène inférieur du Bassin de Paris, 135-194, 4 fig., 2 tabl., 10 pl. VOLUME 2 Fasc. 1 (1968) M. HUGUENEY.— Les gliridés (Rodentia) de l'Oligocène supérieur de St-Victor-la-Coste (Gard), 1-16, 3 pl. Fasc. 2 (1969) J.E. GUILDAY, H.W. HAMILTON & A.D. McCRADY.— Pleistocene vertebrate fauna of Robinson Cave, Overton County, Tennessee, 25-75, 15 fig., 28 tabl. J. de PORTA.— Les vertébrés fossiles de Colombie et les problèmes posés par l'isolement du continent sud-américain, 77-94, 2 fig. Fasc. 3 (1969) J. SUDRE.— Les gisements de Robiac (Eocène supérieur) et leurs faunes de mammifères, 95-156, 21 fig., 5 tabl. Fasc. 4 (1969) M.R. DAWSON.— Osteology of Prolagus sardus, a Quaternary ochotonid (Mammalia, Lagomorpha), 157-190, 38 fig., 1 tabl., 1 pl. Fasc. 5 (1969) L. THALER.— Rongeurs nouveaux de l'Oligocène moyen d'Espagne, 191-207, 9 fig. M. VIANEY-LIAUD.— Rongeurs de l'Oligocène moyen provenant de nouvelles fouilles dans les Phosphorites du Quercy, 209-239, 16 fig. -

Problems of Classification As Applied to the Rodentia
263 PROBLEMS OF CLASSIFICATION AS APPLIED TO THE RODENTIA by Albert E. WOOD* ABSTRACT A classification should be both usable and useful, not too complex either in the amount of splitting or in the number of hierarchies involved, and not so simple as to give a false assurance of knowledge of relationships. Classifi cations are only possible because we do not have complete knowledge of the evolution of the organisms concerned, because gaps in the record are necessary to allow the separation of the various taxa. Rodent classification is compli cated by the large number of organisms involved and by the great amount of parallelism that has taken place In the evolution of any and all features. If several independent features are characteristic of a certain taxon, should an effort be made to define the group on the basis of all the features, or should only one be selected as the determi nant ? Unless the evolution of the several features was closely linked, the former solution will sooner or later lead to insurmountable problems. A classification is a formal arrangement that expresses the author's opinion of the relationships of the organisms concerned. It should be an attempt to approximate the actual genetic relationships existing, or that formerly existed, among the pertinent organisms. During the course of organizing a classification of the mammalian Order Rodentia, I encountered a number of problems of a general nature, some of which are discussed below. Usually, there are extensive gaps in our knowledge of organisms, particularly of fossil ones. These are useful in classification, because we use the gaps to delimit the various units being classified. -

Palaeogene Fossil Record, Phylogeny, Dental Evolution and Historical Biogeography Laurent Marivaux, Myriam Boivin
Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution and historical biogeography Laurent Marivaux, Myriam Boivin To cite this version: Laurent Marivaux, Myriam Boivin. Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution and historical biogeography. Zoological Journal of the Linnean Society, Linnean Society of London, 2019, 187 (3), pp.929-964. 10.1093/zoolinnean/zlz048. hal-02263901 HAL Id: hal-02263901 https://hal.umontpellier.fr/hal-02263901 Submitted on 1 Nov 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Zoological Journal of the Linnean Society Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution, and historical biogeography Journal: Zoological Journal of the Linnean Society Manuscript ID ZOJ-12-2018-3565.R2 Manuscript Type:ForOriginal Review Article Only Africa < Geography, Asia < Geography, mandible < Anatomy, dental Keywords: evolution < Evolution, Eocene < Palaeontology, Oligocene < Palaeontology Although phylogenies imply Asia as the ancestral homeland of the Hystricognathi clade, curiously the oldest known fossil occurrences are not from Asia but from Africa and South America, where they appear by the late middle Eocene. Here we performed cladistic and Bayesian assessments of the dental evidence documenting early ctenohystricans, including several Asian “ctenodactyloids”, all Palaeogene Asian and African hystricognaths, and two early South American hystricognaths. -

Knowledge of the Evolution of African Paleogene Mammals
Knowledge of the évolution of African Paleogene mammals Contribution of the Bir El Ater locality (Eocène, Algeria) Rodolphe Tabuce Brigitte Coiffait Philippe-Emmanuel Coiffait Mohamed Mahboubi Jean-Jacques Jaeger 1 1 Introduction: The Early African Paleogene mammals The African fossil record of therians begins with the Early Cretaceous ofthe Middle Atlas (Morocco) (Sigogneau-Russell, 1991); however, the modem mammalian orders appear only during the early Tertiary in North Africa (figure 1A). The Paleocene and Ypresian localities from the Ouarzazate Basin (Morocco) hâve yielded mammalian faunas with possible creodonts, « insectivores » (paleoryctids, todralestids and adapisoriculids) (Gheerbrant, 1992, 1994, 1995), the oldest représentative ofeupri- mates (Sigé et al, 1990) and archaic ungulates (Sudre et al, 1993). Recently, the discovery of Phosphatherium (Proboscidea) in J\) .... Ypresian Early to Mlddla Mlddlalo Lata C> mlcldla BOCena eocena Ialaeocene eocena "III N'Tagourt2 ElKohol GllbZagdou GourLazib Chambl lnTaliclel M'Bodlone Bir el Ater DoralTalha Qaar (Moroc:co) (Algerla) (Algerla) (Algerla) (Tunlala) and (Sanagal) (Algarla) ·Evaporlt e1Sagha lamagullell Unit· (Fayum, (Mali) (LIbye) Egypt) Matatharla Kassarinolherium tunisiansa Creodonta Koholis gen. el sp. Aprarodon stlasansa indet Hyaanodon Carnlvora Glibzagdouis I8balbslsensis Condylarthra Condyfarthra Condylarthra Inda!. Inda!. Artlodaetyla cl.Bolhrio- ganys sp. Proboscldell KhSmsBcornJS Numidotherium Moerilharium Moenlherium Moarilherium 8srytherium 8srythenum buJbosus -
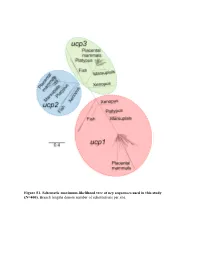
Figure S1. Schematic Maximum-Likelihood Tree of Ucp Sequences Used in This Study (N=400)
Figure S1. Schematic maximum-likelihood tree of ucp sequences used in this study (N=400). Branch lengths denote number of substitutions per site. Exon 1 1 10 20 30 40 50 |...|....|....|....|....|....|....|....|....|....| Homo sapiens ATGGGGGGCCTGA-CAGCCTCGGACGTACACCCGACCC---TGGGGGTCC Bradypus variegatus ?????????????????????????????????????????????????? Choloepus hoffmanni ?????????????????????????????????????????????????? Mylodon darwinii ?????????????????????????????????????????????????? Cyclopes didactylus ?????????????????????????????????????????????????? Dasypus novemcinctus ATGGGGCGCCAGGGCTCCCGCGGGCTCACCCCCCGC-------------- Procavia capensis CTG--AGTTAAGA-CAACCTCAGAAATGCCGCCTAC-----------GCA Elephas maximus ACGGTAGGCCAGA-CGACCGCAGACGTGCCCCGGACCATGGTGGGGGTCA Mammuthus primigenius ACGGTAGGCCAGA-CGACCGCAGACGTGCCCCGGACCATGGTGGGGGTCA Loxodonta africana ACCGTAGGCCAGA-CGACCGCAGACGTGCCCCGGACCATGGTGGGGGTCA Trichechus manatus ATGGTGGGCCAGA-CTACCTGGGATGTGCCCCCGACCA---TGGGCGTCA Dugong dugon ATGGTGGGCCAGA-CTACCTCGGATGTGCCCCCGACCA---TGGGCGTCA Hydrodamalis gigas ATGGTGGGCCAGA-CTACCTCGGATGTGCCCCCGACCA---TGGGCGTCA Sus scrofa CTGTCAGGA-TGA-CAGTTCCTGAAGTGCCCCCGACCA---TAGCGGTCA Sus verrucosus CTGTCAGGA-TGA-CAGTTCCTGAAGTGCCCTCGACCA---TAGCGGTCA Sus cebifrons CTGTCAGGA-TGA-CAGTTCCTGAAGTGCCCTCGACCA---TAGCGGTCA Physeter macrocephalus ATGGTGGGACTCG-CAGCCTCATACGTGCCCCCGACCA---TGGCGGTCA Delphinapterus leucas -------------------------------------------------- Lipotes vexillifer ATGGTGGGACTCG-CAGCCTCAGACGTGCCCCCGACCA---TGGCGGTCA Balaena mysticetus ATGGTGGCACTCA-CAGCCTCAGACGTGCCCCCGACCA---TGGCGGTCA -

Aardvark 25, 53 Abalosia 146 Abrocoma 141, 386 Abrocomidae 29
Cambridge University Press 978-1-107-04433-3 -Evolution of the Rodents: Advances in Phylogeny, Functional Morphology and Development Edited by Philip G. Cox and Lionel Hautier Index More information Index aardvark 25, 53 Alagomyidae 74–5 Abalosia 146 Alagomyidea 75, 81 Abrocoma 141, 386 Alagomys 73–5 Abrocomidae 29, 141, 283, 483 dentition 75 Abudhabia 195 Alagomys inopinatus 74 Acarechimys 146 Alagomys oriensis 74 Acaremyidae 406 Alagomys russelli 74 Acaremys 146 Allactaga 388, 397 Acomys 188–91, 193, 203, 391 Allactaginae 43, 388 Acomys cahirinus 203 allometry 235, 285, 287, 311, 314, 533 Acomys chrysophilus 203 and molars 315 Acomys coppensi 192 Allophaiomys 495, 498 Acomys ineptus 203 Allotheria 324 Acomys kaiseri 203 Alphagaulus 431 Acomys lavocati 203 Anagalida 5–6 Acomys spinosissimus 190–1 anagenesis 140, 150, 154–6 Acomys subspinosus 190 Andinomys 466 Aconaemys porteri 144 annular ligament 382, 389 Aconaemys sagei 144 anoles 57 acouchis 29 Anomaluridae 26, 43, 81, 387–8, 435, 483 acoustic impedance 394 Anomaluroidea 21, 71, 378, 383, 387, 393 acoustic stiffness 394 middle ear 387 Actenomys 148 origin and dispersal 100 Actenomys priscus 144, 150 ossicles 392 adaptive radiation 55–6, 100, 202, 277, 539 Anomaluromorpha 26–8, 43, 290–1, 426, 483 Palaeogene Asian radiation 98 Anomalurus 26, 388 placental 56 anteaters 25, 53 adelomyine 393 Antemus 51, 193, 195 Adelphomys 145 Antemus chinjiensis 192–3, 196 Adu Asa (fossil site) 199 Antemus mancharensis 51, 192–3 Aeretes 223 Anthropoidea 101, 328 Aeromys thomasi 338–9 Apatemyidae 3 Aethomys -

A Fossil History of Southern African Land Mammals
Downloaded from https://www.cambridge.org/core. IP address: 170.106.40.139, on 29 Sep 2021 at 12:08:23, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms. https://www.cambridge.org/core/product/41969EC1E7739F4775954E6ADA8EA036 Downloaded from https://www.cambridge.org/core. IP address: 170.106.40.139, on 29 Sep 2021 at 12:08:23, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms. https://www.cambridge.org/core/product/41969EC1E7739F4775954E6ADA8EA036 A Fossil History of Southern African Land Mammals There is an ever-growing wealth of mammalian fossil material being collected from palaeontological and archaeological sites in southern Africa. This reference provides comprehensive information on the taxonomy and distribution in time and space of all currently recognised southern African fossil mammals. After an introductory background chapter on southern Africa, mammals, sites and dating, the following chapters are presented by epoch, covering the Eocene, Miocene, Pliocene, Pleistocene and Holocene. Individual maps provide information on where in the landscape specific taxa have been found, and a comprehensive index lists all the fauna and site locations. It ends with a chapter on how the book can be used, and lines of future research. Collecting a vast amount of information together in an accessible format, this is an essential reference for non-specialist taxonomists and palaeontologists, as well as for those using fossil data for other applications, such as archaeology, neontology and nature conservation. This title is also available as Open Access on Cambridge Core. D. Margaret Avery is Emeritus Associate of Cenozoic Studies at Iziko Museums of South Africa, and Honorary Researcher at the Evolutionary Studies Institute at the University of Witwatersrand. -

Fossil and Molecular Evidence Constrain Scenarios for the Early Evolutionary and Biogeographic History of Hystricognathous Rodents
Fossil and molecular evidence constrain scenarios for the early evolutionary and biogeographic history of hystricognathous rodents Hesham M. Sallama,1, Erik R. Seiffertb, Michael E. Steiperc,d, and Elwyn L. Simonse;1 aDepartment of Earth Sciences, University of Oxford, Parks Road, Oxford OX1 3PR, United Kingdom; bDepartment of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794; cDepartment of Anthropology, Hunter College of the City University of New York, 695 Park Avenue, NY 10065; dDepartments of Anthropology and Biology, Graduate Center of the City University of New York, 365 Fifth Avenue, NY 10016; and eDivision of Fossil Primates, Duke Lemur Center, 1013 Broad Street, Durham, NC 27705 Contributed by Elwyn L. Simons, Duke Lemur Center, Durham, NC, August 11, 2009 (sent for review July 12, 2009) The early evolutionary and paleobiogeographic history of the diverse Afro-Arabian and South American distributions, respectively, the early rodent clade Hystricognathi, which contains Hystricidae (Old World biogeographic history of crown Hystricognathi remains a matter of porcupines), Caviomorpha (the endemic South American rodents), persistent debate. Some have argued that phiomorphs and caviomorphs and African Phiomorpha (cane rats, dassie rats, and blesmols) is of are likely to have shared a common Afro-Arabian ancestor (15–17), great interest to students of mammalian evolution, but remains while others have suggested that the phiomorph-caviomorph split might poorly understood because of a poor early fossil record. Here we have occurred in Asia, and that caviomorphs dispersed to South describe the oldest well-dated hystricognathous rodents from an America either through Afro-Arabia or via a southern Gondwanan earliest late Eocene (Ϸ37 Ma) fossil locality in the Fayum Depression route (13, 18). -

Supplementary Materials For
Supplementary Materials for Inactivation of thermogenic UCP1 as a historical contingency in multiple placental mammal clades Michael J. Gaudry, Martin Jastroch, Jason R. Treberg, Michael Hofreiter, Johanna L.A. Paijmans, James Starrett, Nathan Wales, Anthony V. Signore, Mark S. Springer and Kevin L. Campbell correspondence to: [email protected] This PDF file includes: Materials and Methods Figs. S1 to S4 Tables S1 to S7 References Other Supplementary Materials for this manuscript include the following: Data files S1 to S10 Data file S1: ucp1 free ratio model (Text file) Data file S2: gene partitions, a maximum likelihood phylogram, and mcmctree (Text file) Data file S3: ucp2 free ratio model (Text file) Data file S4: ucp3 free ratio model (Text file) Data file S5: ucp1 alignment (Fasta file) Data file S6: ucp2 alignment (Fasta file) Data file S7: ucp3 alignment (Fasta file) Data file S8: Accesion numbers for the 41 loci used to contruct the 51 kb maximum likelihood species tree (Fasta file) Data file S9: 51 kb alignment (41 genes) for 140 vertebrate species (Nexus file) Data file S10: Fossil body mass, taxon diversity, and UCP1 diversification codings (Excel file) Materials and Methods Taxon Sampling. This study included representatives of 141 vertebrates, 133 of which are mammals (1 monotreme, 4 marsupials, 4 xenarthrans, 11 afrotherians, 59 laurasiatherians, and 54 euarchontoglires). Outgroup representatives included 7 fish and 1 amphibian species. Due to the documented loss of uncoupling protein 1 (ucp1) in the archosaur lineage (55), birds and reptiles were not included in the dataset. We employed genome mining, polymerase chain reaction (PCR), and hybridization capture to obtain ucp1, ucp2, ucp3, and sarcolipin (sln) coding sequences examined in this study. -

GSN Comms 19. 7. Pickford, Tufamys
Tufamyidae, a new family of hystricognath rodents from the Palaeogene and Neogene of the Sperrgebiet, Namibia Martin PICKFORD Sorbonne Universités (CR2P, MNHN, CNRS, UPMC - Paris VI) 8, rue Buffon, 75005, Paris, France, e-mail : [email protected] Abstract : The Palaeogene tufa deposits at Eocliff are a rich source of micromammals, probably the richest such site in Africa, having yielded thousands of specimens, including complete skulls, mandibles and associated post-cranial elements, with perhaps millions of specimens remaining in the field. Rodents are well-represented but are of relatively low diversity, comprising six common species and a rare one. We here describe an early hystricognath, hystricomorph rodent from the site, based on a complete skull associated with post-cranial elements, and many maxillae, mandibles, isolated teeth and post-cranial bones. This is currently the earliest known member of the superfamily Thryonomyoidea, possible earlier records being based on isolated teeth or fragmentary maxillae and mandibles which do not provide solid evidence for their superfamilial status. The originality of the morphology of the incisors, the simplicity of the molars and other features indicate that this thryonomyoid represents a hitherto unknown family of rodents, herein named Tufamyidae. Key words : Eocene; Biogeography; Evolution; Taxonomy; Systematics, Rodentia. To cite this paper : Pickford, M. 2018. Tufamyidae, a new family of hystricognath rodents from the Palaeogene and Neogene of the Sperrgebiet, Namibia. Communications of the Geological Survey of Namibia , 19: 71-109. Introduction The tufa deposits at Eocliff and Eoridge in This article focusses on one of the common the Sperrgebiet, Namibia, have yielded an rodent species at Eocliff, being the first of a impressive diversity of micromammals of Late series of papers treating each of the genera Eocene age, among which rodents are occurring at the site. -

American Museum Novitates
AMERICAN MUSEUM NOVITATES Number 3750, 36 pp. July 20, 2012 Two New Taxa (Caviomorpha, Rodentia) from the Early Oligocene Tinguiririca Fauna (Chile) ORNELLA C. BERTRAND,1 JOHN J. FLYNN,2 DARIN A. CROFT,3 AND ANDRE R. WYSS4 ABSTRACT Here we describe two new caviomorphs from the early Oligocene Tinguiririca Fauna of the Andean Main Range of central Chile, representing the most ancient rodents known from the mid to high latitudes of South America, and the second-oldest securely dated rodents from the continent. These two new taxa are each documented by single partial mandibles bearing largely complete dentitions. Representing two new taxa, Andemys termasi, gen. et sp. nov., and Eoviscaccia frassinettii, sp. nov., these caviomorphs are informally referred to the pan-Dasy- proctidae and pan-Chinchillidae, respectively. These taxa, together with recent findings in Peru, confirm that caviomorphs were well diversified prior to the Deseadan SALMA, that they likely originated during the middle to late Eocene, but that they did not spread from the tropics until some time after the Mustersan—a well-sampled interval from which rodents are unknown in higher latitudes. Additionally, in documenting the earliest occurrence of hypsodonty among caviomorphs Eoviscaccia frassinettii, sp. nov., provides important insights into the acquisition of this common mammalian dental innovation in rodents. 1 Laboratoire de Paléontologie, Institut des Sciences de l’Évolution, c.c. 64, Université Montpellier II, Place Eugène Bataillon, F-34095 Montpellier cedex 05, France; and Kade Fellow, Richard Gilder Graduate School, American Museum of Natural History, Central Park West at 79th St., New York, NY 10024. 2 Division of Paleontology and Richard Gilder Graduate School, American Museum of Natural History.