Endorsing Darwin – Global Biogeography of the Epipelagic Goose Barnacles Lepas Spp
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
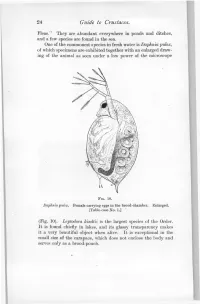
24 Guide to Crustacea
24 Guide to Crustacea. Fleas." They are abundant everywhere in ponds and ditches, and a few species are found in the sea. One of the commonest species in fresh water is Daphnia pulex, of which specimens are exhibited together with an enlarged draw- ing of the animal as seen under a low power of the microscope FIG. 10. Daphnia pulex. Female carrying eggs in the brood-chamber. Enlarged. [Table-case No. 1.] (Fig. 10). Leptodora kindtii is the largest species of the Order. It is found chiefly in lakes, and its glassy transparency makes it a very beautiful object when alive. It is exceptional in the small size of the carapace, which does not enclose the body and serves only as a brood-pouch. Ostracoda. 25 Sub-class II.—OSTRACODA. (Table-ease No. 1.) The number of somites, as indicated by the appendages, is smaller than in any other Crustacea, there being, at most, only two pairs of trunk-limbs behind those belonging to the head- region. The carapace forms a bivalved shell completely en- closing the body and limbs. There is a large, and often leg-like, palp on the mandible. The antennules and antennae are used for creeping or swimming. The Ostracoda (Fig. 10) are for the most part extremely minute animals, and only one or two of the larger species can be exhibited. They occur abundantly in fresh water and in FIG. 11. Shells of Ostracoda, seen from the side. A. Philomedes brenda (Myodocopa) ; B. Cypris fuscata (Podocopa); ('. Cythereis ornata (Podocopa): all much enlarged, n., Notch characteristic of the Myodocopa; e., the median eye ; a., mark of attachment of the muscle connecting the two valves of the shell. -

Kinematic and Dynamic Scaling of Copepod Swimming
fluids Review Kinematic and Dynamic Scaling of Copepod Swimming Leonid Svetlichny 1,* , Poul S. Larsen 2 and Thomas Kiørboe 3 1 I.I. Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Str. B. Khmelnytskogo, 15, 01030 Kyiv, Ukraine 2 DTU Mechanical Engineering, Fluid Mechanics, Technical University of Denmark, Building 403, DK-2800 Kgs. Lyngby, Denmark; [email protected] 3 Centre for Ocean Life, Danish Technical University, DTU Aqua, Building 202, DK-2800 Kgs. Lyngby, Denmark; [email protected] * Correspondence: [email protected] Received: 30 March 2020; Accepted: 6 May 2020; Published: 11 May 2020 Abstract: Calanoid copepods have two swimming gaits, namely cruise swimming that is propelled by the beating of the cephalic feeding appendages and short-lasting jumps that are propelled by the power strokes of the four or five pairs of thoracal swimming legs. The latter may be 100 times faster than the former, and the required forces and power production are consequently much larger. Here, we estimated the magnitude and size scaling of swimming speed, leg beat frequency, forces, power requirements, and energetics of these two propulsion modes. We used data from the literature together with new data to estimate forces by two different approaches in 37 species of calanoid copepods: the direct measurement of forces produced by copepods attached to a tensiometer and the indirect estimation of forces from swimming speed or acceleration in combination with experimentally estimated drag coefficients. Depending on the approach, we found that the propulsive forces, both for cruise swimming and escape jumps, scaled with prosome length (L) to a power between 2 and 3. -

Parallel Changes in Genital Morphology Delineate Cryptic Diversification of Planktonic Nudibranchs
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Poet Commons (Whittier College) Whittier College Poet Commons Biology Faculty Publications & Research 2013 Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs Alvin Alejandrino [email protected] Celia K. C. Churchill Ángel Valdés Diarmaid Ó. Foighil Follow this and additional works at: https://poetcommons.whittier.edu/bio Part of the Biology Commons Recommended Citation Alejandrino, Alvin; Churchill, Celia K. C.; Valdés, Ángel; and Foighil, Diarmaid Ó., "Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs" (2013). Biology. 1. https://poetcommons.whittier.edu/bio/1 This Article is brought to you for free and open access by the Faculty Publications & Research at Poet Commons. It has been accepted for inclusion in Biology by an authorized administrator of Poet Commons. For more information, please contact [email protected]. Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs rspb.royalsocietypublishing.org Celia K. C. Churchill1,†, Alvin Alejandrino2,‡,A´ngel Valde´s2 and Diarmaid O´ Foighil1 1Museum of Zoology, Department of Ecology and Evolutionary Biology, University of Michigan, 1109 Geddes Avenue, Ann Arbor, MI 48109-1079, USA 2Department of Biological Sciences, California State Polytechnic University, 3801 West Temple Avenue, Pomona, Research CA 91768, USA Cite this article: Churchill CKC, Alejandrino A, The relative roles of geographical and non-geographical barriers in the genesis Valde´sA´,O´ Foighil D. 2013 Parallel changes in of genetic isolation are highly debated in evolutionary biology, yet knowing genital morphology delineate cryptic diversifi- how speciation occurs is essential to our understanding of biodiversity. -

Darwin in the Press: What the Spanish Dailies Said About the 200Th Anniversary of Charles Darwin’S Birth
CONTRIBUTIONS to SCIENCE, 5 (2): 193–198 (2009) Institut d’Estudis Catalans, Barcelona Celebration of the Darwin Year 2009 DOI: 10.2436/20.7010.01.75 ISSN: 1575-6343 www.cat-science.cat Darwin in the press: What the Spanish dailies said about the 200th anniversary of Charles Darwin’s birth Esther Díez, Anna Mateu, Martí Domínguez Mètode, University of Valencia, Valencia, Spain Resum. El bicentenari del naixement de Charles Darwin, cele- Summary. The bicentenary of Charles Darwin’s birth, cele- brat durant el passat any 2009, va provocar un considerable brated in 2009, caused a surge in the interest shown by the augment de l’interés informatiu de la figura del naturalista an- press in the life and work of the renowned English naturalist. glès. En aquest article s’analitza la cobertura informativa This article analyzes the press coverage devoted to the Darwin d’aquest aniversari en onze diaris generalistes espanyols, i Year in eleven Spanish daily papers during the week of Dar- s’estableix grosso modo quines són les principals postures de win’s birthday and provides a brief synopsis of the positions la premsa espanyola davant de la teoria de l’evolució. D’aques- adopted by the Spanish press regarding the theory of evolu- ta anàlisi, queda ben palès com el creacionisme és encara molt tion. The analysis makes very clear the relatively strong support viu en els mitjans espanyols més conservadors. for creationism by the conservative press in Spain. Paraules clau: Charles Darwin ∙ ciència vs. creacionisme ∙ Keywords: Charles Darwin ∙ science vs. creationism ∙ tractament informatiu científic scientific news coverage “The more we know of the fixed laws of nature the more In this article, we examine to what extent and in what form incredible do miracles become.” this controversy persists in Spanish society, focusing on the print media’s response to the celebration, in 2009, of the 200th Charles Darwin (1887). -

Xoimi AMERICAN COXCIIOLOGY
S31ITnS0NIAN MISCEllANEOUS COLLECTIOXS. BIBLIOGIIAPHY XOimi AMERICAN COXCIIOLOGY TREVIOUS TO THE YEAR 18G0. PREPARED FOR THE SMITHSONIAN INSTITUTION BY . W. G. BINNEY. PART II. FOKEIGN AUTHORS. WASHINGTON: SMITHSONIAN INSTITUTION. JUNE, 1864. : ADYERTISEMENT, The first part of the Bibliography of American Conchology, prepared for the Smithsonian Institution by Mr. Binuey, was published in March, 1863, and embraced the references to de- scriptions of shells by American authors. The second part of the same work is herewith presented to the public, and relates to species of North American shells referred to by European authors. In foreign works binomial authors alone have been quoted, and no species mentioned which is not referred to North America or some specified locality of it. The third part (in an advanced stage of preparation) will in- clude the General Index of Authors, the Index of Generic and Specific names, and a History of American Conchology, together with any additional references belonging to Part I and II, that may be met with. JOSEPH HENRY, Secretary S. I. Washington, June, 1864. (" ) PHILADELPHIA COLLINS, PRINTER. CO]^TENTS. Advertisement ii 4 PART II.—FOREIGN AUTHORS. Titles of Works and Articles published by Foreign Authors . 1 Appendix II to Part I, Section A 271 Appendix III to Part I, Section C 281 287 Appendix IV .......... • Index of Authors in Part II 295 Errata ' 306 (iii ) PART II. FOEEIGN AUTHORS. ( V ) BIBLIOGRxVPHY NOETH AMERICAN CONCHOLOGY. PART II. Pllipps.—A Voyage towards the North Pole, &c. : by CON- STANTiNE John Phipps. Loudou, ITTJc. Pa. BIBLIOGRAPHY OF [part II. FaliricillS.—Fauna Grcenlandica—systematice sistens ani- malia GrcEulandite occidentalis liactenus iudagata, &c., secun dum proprias observatioues Othonis Fabricii. -

The XMM-Newton SAS - Distributed Development and Maintenance of a Large Science Analysis System: a Critical Analysis
Astronomical Data Analysis Software and Systems XIII ASP Conference Series, Vol. 314, 2004 F. Ochsenbein, M. Allen, and D. Egret, eds. The XMM-Newton SAS - Distributed Development and Maintenance of a Large Science Analysis System: A Critical Analysis Carlos Gabriel1, John Hoar1, Aitor Ibarra1, Uwe Lammers2, Eduardo Ojero1, Richard Saxton1, Giuseppe Vacanti2 XMM-Newton Science Operations Centre, Science Operations and Data Systems Division of ESA, European Space Agency Mike Denby, Duncan Fyfe, Julian Osborne Department of Physics and Astronomy, University of Leicester, Leicester, LE1 7RH, UK Abstract. The XMM-Newton Scientific Analysis System (SAS) is the software used for the reduction and calibration of data taken with the XMM-Newton satellite instruments leading to almost 400 refereed scien- tific papers published in the last 2.5 years. Its maintenance, further devel- opment and distribution is under the responsibility of the XMM-Newton Science Operations Centre together with the Survey Science Centre, rep- resenting a collaborative effort of more than 30 scientific institutes. Developed in C++, Fortran 90/95 and Perl, the SAS makes large use of open software packages such as ds9 for image display (SAO-R&D Software Suite), Grace, LHEASOFT and cfitsio (HEASARC project), pgplot, fftw and the non-commercial version of Qt (TrollTech). The combination of supporting several versions of SAS for multiple platforms (including SunOS, DEC, many Linux flavours and MacOS) in a widely distributed development process which makes use of a suite of ex- ternal packages and libraries presents substantial issues for the integrity of the SAS maintenance and development. A further challenge comes from the necessity of maintaining the flexibility of a software package evolving together with progress made in instrument calibration and anal- ysis refinement, whilst at the same time being the source of all official products of the XMM-Newton mission. -

Tesis Estructura Comunitaria De Copepodos .Pdf
Universidad de Concepción Dirección de Postgrado Facultad de Ciencias Naturales y Oceanográficas Programa de Magister en Ciencias mención Oceanografía Estructura comunitaria de copépodos pelágicos asociados a montes submarinos de la Dorsal Juan Fernández (32-34°S) en el Pacífico Sur Oriental Tesis para optar al grado de Magíster en Ciencias con mención en Oceanografía PAMELA ANDREA FIERRO GONZÁLEZ CONCEPCIÓN-CHILE 2019 Profesora Guía: Pamela Hidalgo Díaz Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas Universidad de Concepción Profesor Co-guía: Rubén Escribano Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas Universidad de Concepción La Tesis de “Magister en Ciencias con mención en Oceanografía” titulada “Estructura comunitaria de copépodos pelágicos asociados a montes submarinos de la Dorsal Juan Fernández (32-34°S) en el Pacífico sur oriental”, de la Srta. “PAMELA ANDREA FIERRO GONZÁLEZ” y realizada bajo la Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, ha sido aprobada por la siguiente Comisión de Evaluación: Dra. Pamela Hidalgo Díaz Profesora Guía Universidad de Concepción Dr. Rubén Escribano Profesor Co-Guía Universidad de Concepción Dr. Samuel Hormazábal Miembro de la Comisión Evaluadora Pontificia Universidad Católica de Valparaíso Dr. Fabián Tapia Director Programa de Magister en Oceanografía Universidad de Concepción ii A Juan Carlos y Sebastián iii AGRADECIMIENTOS Agradezco a quienes con su colaboración y apoyo hicieron posible el desarrollo y término de esta tesis. En primer lugar, agradezco a los miembros de mi comisión de tesis. A mi profesora guía, Dra. Pamela Hidalgo, por apoyarme y guiarme en este largo camino de formación académica, por su gran calidad humana, contención y apoyo personal. -

Prime Focus (06-16).Pub
Highlights of the June Sky - - - - - - Saturn at opposition. - - - - - - DAWN: A very thin waning A Publication of the Kalamazoo Astronomical Society crescent Moon is about 2° below Mercury. Look low in east about 20 minutes before sunrise. - - - 4th - - - New Moon 11:00 pm EDT KAS - - - 9th - - - PM: Regulus, the brightest star in Leo, is about 7° to General Meeting: Friday, June 3 @ 7:00 pm the Moon’s upper left. Kalamazoo Area Math & Science Center - See Page 8 for Details - - - 10th - - - PM: The Moon is located about halfway between Observing Session: Saturday, June 11 @ 9:30 pm Regulus and Jupiter. The Moon, Mars, Jupiter & Saturn - Kalamazoo Nature Center - - - 11th - - - PM: The Moon forms a Board Meeting: Sunday, June 12 @ 5:00 pm triangle with Jupiter and Sunnyside Church - 2800 Gull Road - All Members Welcome Sigma () Leonis. - - - 12th - - - Observing Session: Saturday, June 25 @ 9:30 pm First Quarter Moon 4:10 am EDT Grand Globular Clusters - Kalamazoo Nature Center - - - 14th - - - PM: Spica, the brightest star in Virgo, is <5° below a waxing gibbous Moon. - - - 17th → 18th - - - Inside the Newsletter. PM: The Moon, Saturn, and Mars make a wide, flat May Meeting Minutes............................. p. 2 triangle, with the longest side stretching ~18° to Board Meeting Minutes......................... p. 3 connect the two planets. - - - 18th → 19th - - - Learn More About the A.L...................p. 3 PM: The Moon and Saturn are 3° to 4° apart. Observations of the Red Planet.......... p. 4 - - - 20th - - - NASA Space Place.................................. p. 5 Full Moon 7:02 am EDT June Night Sky......................................... p. 6 Summer solstice occurs at KAS Board & Announcements........... -

A Checklist of Turtle and Whale Barnacles
Journal of the Marine Biological Association of the United Kingdom, 2013, 93(1), 143–182. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S0025315412000847 A checklist of turtle and whale barnacles (Cirripedia: Thoracica: Coronuloidea) ryota hayashi1,2 1International Coastal Research Center, Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5, Kashiwanoha, Kashiwa-shi, Chiba 277-8564 Japan, 2Marine Biology and Ecology Research Program, Extremobiosphere Research Center, Japan Agency for Marine–Earth Science and Technology A checklist of published records of coronuloid barnacles (Cirripedia: Thoracica: Coronuloidea) attached to marine vertebrates is presented, with 44 species (including 15 fossil species) belonging to 14 genera (including 3 fossil genera) and 3 families recorded. Also included is information on their geographical distribution and the hosts with which they occur. Keywords: checklist, turtle barnacles, whale barnacles, Chelonibiidae, Emersoniidae, Coronulidae, Platylepadidae, host and distribution Submitted 10 May 2012; accepted 16 May 2012; first published online 10 August 2012 INTRODUCTION Superorder THORACICA Darwin, 1854 Order SESSILIA Lamarck, 1818 In this paper, a checklist of barnacles of the superfamily Suborder BALANOMORPHA Pilsbry, 1916 Coronuloidea occurring on marine animals is presented. Superfamily CORONULOIDEA Newman & Ross, 1976 The systematic arrangement used herein follows Newman Family CHELONIBIIDAE Pilsbry, 1916 (1996) rather than Ross & Frick (2011) for reasons taken up in Hayashi (2012) in some detail. The present author Genus Chelonibia Leach, 1817 deems the subfamilies of the Cheonibiidae (Chelonibiinae, Chelonibia caretta (Spengler, 1790) Emersoniinae and Protochelonibiinae) proposed by Harzhauser et al. (2011), as well as those included of Ross & Lepas caretta Spengler, 1790: 185, plate 6, figure 5. -

Cirripedia of Madeira
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Universidade do Algarve Helgol Mar Res (2006) 60: 207–212 DOI 10.1007/s10152-006-0036-5 ORIGINAL ARTICLE Peter Wirtz Æ Ricardo Arau´jo Æ Alan J. Southward Cirripedia of Madeira Received: 13 September 2005 / Revised: 12 January 2006 / Accepted: 13 January 2006 / Published online: 3 February 2006 Ó Springer-Verlag and AWI 2006 Abstract We give a list of Cirripedia from Madeira phers. The marine invertebrates have been less studied Island and nearby deep water, based on specimens in and there has been no compilation of cirripede records the collection of the Museu Municipal do Funchal for Madeira, comparable to those for the Azores (Histo´ria Natural) (MMF), records mentioned in the archipelago (Young 1998a; Southward 1999). We here literature, and recent collections. Tesseropora atlantica summarize records from Madeira and nearby deep water Newman and Ross, 1976 is recorded from Madeira for and discuss their biogeographical implications. the first time. The Megabalanus of Madeira is M. az- oricus. There are 20 genera containing 27 species, of which 22 occur in depths less than 200 m. Of these Methods shallow water species, eight are wide-ranging oceanic forms that attach to other organisms or to floating The records are based on (1) the work of R.T. Lowe, objects, leaving just 13 truly benthic shallow water who sent specimens to Charles Darwin; (2) material in barnacles. This low diversity is probably a consequence the Museu Municipal do Funchal (Histo´ria Natural) of the distance from the continental coasts and the (MMF); (3) casual collecting carried out by residents or small area of the available habitat. -

SUCCESSES of the HUBBLE SPACE TELESCOPE, 18Th February 2005 Astronomical History the Hubble Telescope Has Been a Major Tool for the Last Fifteen Years
SUCCESSES OF THE HUBBLE SPACE TELESCOPE, 18th February 2005 Astronomical History The Hubble telescope has been a major tool for the last fifteen years. However, astronomy is as old as man – the earliest known good star map was by Hipparchus 2500 years ago, and a Roman statue (now in Naples) is of Atlas supporting a celestial globe. In 1610 Galileo made the first telescope, opening up the sky; he was able to see the moons of Jupiter, which clearly orbited Jupiter – not Earth, then considered the absolute centre of the universe… Larger telescopes were soon built, with long tubes to accommodate the low curvature lenses that gave the best results – they were not easily manoeuvred. There is a large 28” Refractor at the Observatory in Greenwich, designated ROG, made in 1893 but on an 1859 mount, which even in London’s atmosphere is useful for planetary observation. In 19thC astronomers began to study not just the location of celestial objects but what types they were. Reflecting Telescopes These use a mirror as the prime element to gather and focus light. Small ones were made in the late 18thC, then in the early 19thC Herschel designed one with an 18” mirror. Mirrors being supported from behind, are easier to mount than lenses can only be held at the edge; they can gather more light and give a brighter, sharper image. In 1948 the 200” (5 m) Hale telescope at Mount Palomar set a new standard, high up above much of the atmosphere for maximum clarity. This was built for traditional observation by eye; then longer exposures were made possible with photographic plates; now CCDs (Charge Coupled Detectors) give higher sensitivity. -

A Comparison of Copepoda (Order: Calanoida, Cyclopoida, Poecilostomatoida) Density in the Florida Current Off Fort Lauderdale, Florida
Nova Southeastern University NSUWorks HCNSO Student Theses and Dissertations HCNSO Student Work 6-1-2010 A Comparison of Copepoda (Order: Calanoida, Cyclopoida, Poecilostomatoida) Density in the Florida Current Off orF t Lauderdale, Florida Jessica L. Bostock Nova Southeastern University, [email protected] Follow this and additional works at: https://nsuworks.nova.edu/occ_stuetd Part of the Marine Biology Commons, and the Oceanography and Atmospheric Sciences and Meteorology Commons Share Feedback About This Item NSUWorks Citation Jessica L. Bostock. 2010. A Comparison of Copepoda (Order: Calanoida, Cyclopoida, Poecilostomatoida) Density in the Florida Current Off Fort Lauderdale, Florida. Master's thesis. Nova Southeastern University. Retrieved from NSUWorks, Oceanographic Center. (92) https://nsuworks.nova.edu/occ_stuetd/92. This Thesis is brought to you by the HCNSO Student Work at NSUWorks. It has been accepted for inclusion in HCNSO Student Theses and Dissertations by an authorized administrator of NSUWorks. For more information, please contact [email protected]. Nova Southeastern University Oceanographic Center A Comparison of Copepoda (Order: Calanoida, Cyclopoida, Poecilostomatoida) Density in the Florida Current off Fort Lauderdale, Florida By Jessica L. Bostock Submitted to the Faculty of Nova Southeastern University Oceanographic Center in partial fulfillment of the requirements for the degree of Master of Science with a specialty in: Marine Biology Nova Southeastern University June 2010 1 Thesis of Jessica L. Bostock Submitted in Partial Fulfillment of the Requirements for the Degree of Masters of Science: Marine Biology Nova Southeastern University Oceanographic Center June 2010 Approved: Thesis Committee Major Professor :______________________________ Amy C. Hirons, Ph.D. Committee Member :___________________________ Alexander Soloviev, Ph.D.