Understanding the Ecology of Ross River Virus; Novel Approaches and Insights Into Non-Human Reservoirs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Climate Change and Human Health: Risks and Responses
Climate change and human health RISKS AND RESPONSES Editors A.J. McMichael The Australian National University, Canberra, Australia D.H. Campbell-Lendrum London School of Hygiene and Tropical Medicine, London, United Kingdom C.F. Corvalán World Health Organization, Geneva, Switzerland K.L. Ebi World Health Organization Regional Office for Europe, European Centre for Environment and Health, Rome, Italy A.K. Githeko Kenya Medical Research Institute, Kisumu, Kenya J.D. Scheraga US Environmental Protection Agency, Washington, DC, USA A. Woodward University of Otago, Wellington, New Zealand WORLD HEALTH ORGANIZATION GENEVA 2003 WHO Library Cataloguing-in-Publication Data Climate change and human health : risks and responses / editors : A. J. McMichael . [et al.] 1.Climate 2.Greenhouse effect 3.Natural disasters 4.Disease transmission 5.Ultraviolet rays—adverse effects 6.Risk assessment I.McMichael, Anthony J. ISBN 92 4 156248 X (NLM classification: WA 30) ©World Health Organization 2003 All rights reserved. Publications of the World Health Organization can be obtained from Marketing and Dis- semination, World Health Organization, 20 Avenue Appia, 1211 Geneva 27, Switzerland (tel: +41 22 791 2476; fax: +41 22 791 4857; email: [email protected]). Requests for permission to reproduce or translate WHO publications—whether for sale or for noncommercial distribution—should be addressed to Publications, at the above address (fax: +41 22 791 4806; email: [email protected]). The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Queensland Public Boat Ramps
Queensland public boat ramps Ramp Location Ramp Location Atherton shire Brisbane city (cont.) Tinaroo (Church Street) Tinaroo Falls Dam Shorncliffe (Jetty Street) Cabbage Tree Creek Boat Harbour—north bank Balonne shire Shorncliffe (Sinbad Street) Cabbage Tree Creek Boat Harbour—north bank St George (Bowen Street) Jack Taylor Weir Shorncliffe (Yundah Street) Cabbage Tree Creek Boat Harbour—north bank Banana shire Wynnum (Glenora Street) Wynnum Creek—north bank Baralaba Weir Dawson River Broadsound shire Callide Dam Biloela—Calvale Road (lower ramp) Carmilla Beach (Carmilla Creek Road) Carmilla Creek—south bank, mouth of creek Callide Dam Biloela—Calvale Road (upper ramp) Clairview Beach (Colonial Drive) Clairview Beach Moura Dawson River—8 km west of Moura St Lawrence (Howards Road– Waverley Creek) Bund Creek—north bank Lake Victoria Callide Creek Bundaberg city Theodore Dawson River Bundaberg (Kirby’s Wall) Burnett River—south bank (5 km east of Bundaberg) Beaudesert shire Bundaberg (Queen Street) Burnett River—north bank (downstream) Logan River (Henderson Street– Henderson Reserve) Logan Reserve Bundaberg (Queen Street) Burnett River—north bank (upstream) Biggenden shire Burdekin shire Paradise Dam–Main Dam 500 m upstream from visitors centre Barramundi Creek (Morris Creek Road) via Hodel Road Boonah shire Cromarty Creek (Boat Ramp Road) via Giru (off the Haughton River) Groper Creek settlement Maroon Dam HG Slatter Park (Hinkson Esplanade) downstream from jetty Moogerah Dam AG Muller Park Groper Creek settlement Bowen shire (Hinkson -

The Non-Human Reservoirs of Ross River Virus: a Systematic Review of the Evidence Eloise B
Stephenson et al. Parasites & Vectors (2018) 11:188 https://doi.org/10.1186/s13071-018-2733-8 REVIEW Open Access The non-human reservoirs of Ross River virus: a systematic review of the evidence Eloise B. Stephenson1*, Alison J. Peel1, Simon A. Reid2, Cassie C. Jansen3,4 and Hamish McCallum1 Abstract: Understanding the non-human reservoirs of zoonotic pathogens is critical for effective disease control, but identifying the relative contributions of the various reservoirs of multi-host pathogens is challenging. For Ross River virus (RRV), knowledge of the transmission dynamics, in particular the role of non-human species, is important. In Australia, RRV accounts for the highest number of human mosquito-borne virus infections. The long held dogma that marsupials are better reservoirs than placental mammals, which are better reservoirs than birds, deserves critical review. We present a review of 50 years of evidence on non-human reservoirs of RRV, which includes experimental infection studies, virus isolation studies and serosurveys. We find that whilst marsupials are competent reservoirs of RRV, there is potential for placental mammals and birds to contribute to transmission dynamics. However, the role of these animals as reservoirs of RRV remains unclear due to fragmented evidence and sampling bias. Future investigations of RRV reservoirs should focus on quantifying complex transmission dynamics across environments. Keywords: Amplifier, Experimental infection, Serology, Virus isolation, Host, Vector-borne disease, Arbovirus Background transmission dynamics among arboviruses has resulted in Vertebrate reservoir hosts multiple definitions for the key term “reservoir” [9]. Given Globally, most pathogens of medical and veterinary im- the diversity of virus-vector-vertebrate host interactions, portance can infect multiple host species [1]. -

Demographic Consequences of Superabundance in Krefft's River
i The comparative ecology of Krefft’s River Turtle Emydura krefftii in Tropical North Queensland. By Dane F. Trembath B.Sc. (Zoology) Applied Ecology Research Group University of Canberra ACT, 2601 Australia A thesis submitted in fulfilment of the requirements of the degree of Masters of Applied Science (Resource Management). August 2005. ii Abstract An ecological study was undertaken on four populations of Krefft’s River Turtle Emydura krefftii inhabiting the Townsville Area of Tropical North Queensland. Two sites were located in the Ross River, which runs through the urban areas of Townsville, and two sites were in rural areas at Alligator Creek and Stuart Creek (known as the Townsville Creeks). Earlier studies of the populations in Ross River had determined that the turtles existed at an exceptionally high density, that is, they were superabundant, and so the Townsville Creek sites were chosen as low abundance sites for comparison. The first aim of this study was to determine if there had been any demographic consequences caused by the abundance of turtle populations of the Ross River. Secondly, the project aimed to determine if the impoundments in the Ross River had affected the freshwater turtle fauna. Specifically this study aimed to determine if there were any difference between the growth, size at maturity, sexual dimorphism, size distribution, and diet of Emydura krefftii inhabiting two very different populations. A mark-recapture program estimated the turtle population sizes at between 490 and 5350 turtles per hectare. Most populations exhibited a predominant female sex-bias over the sampling period. Growth rates were rapid in juveniles but slowed once sexual maturity was attained; in males, growth basically stopped at maturity, but in females, growth continued post-maturity, although at a slower rate. -

Wetlands of the Townsville Area
A Final Report to the Townsville City Council WETLANDS OF THE TOWNSVILLE AREA ACTFR Report 96/28 25 November 1996 Prepared by G. Lukacs of the Australian Centre for Tropical Freshwater Research, James Cook University of North Queensland, Townsville Q 4811 Telephone (077 814262 Facsimile (077) 815589 Wetlands of the TCC LGA: Report No.96/28 TABLE OF CONTENTS 1. INTRODUCTION .................................................................................................................................. 1 1.1 Wetlands and the Community............................................................................................................. 1 1.2 The Wetlands of the Townsville Region ............................................................................................. 1 1.3 Values and Functions of Wetlands..................................................................................................... 3 2. METHODOLOGY ................................................................................................................................. 4 2.1 Scope .................................................................................................................................................. 4 2.2 Mapping ............................................................................................................................................. 4 2.3 Classification ..................................................................................................................................... 5 2.4 Sampling............................................................................................................................................ -

A Short History of Thuringowa
its 0#4, Wdkri Xdor# of fhurrngoraa Published by Thuringowa City Council P.O. Box 86, Thuringowa Central Queensland, 4817 Published October, 2000 Copyright The City of Thuringowa This book is copyright. Apart from any fair dealing for the purposes of private study, research, criticism or review, as permitted under the Copyright Act no part may be reproduced by any process without written permission. Inquiries should be addressed to the Publishers. All rights reserved. ISBN: 0 9577 305 3 5 kk THE CITY of Centenary of Federation i HURINGOWA Queensland This publication is a project initiated and funded by the City of Thuringowa This project is financially assisted by the Queensland Government, through the Queensland Community Assistance Program of the Centenary of Federation Queensland Cover photograph: Ted Gleeson crossing the Bohle. Gleeson Collection, Thuringowa Conienis Forward 5 Setting the Scene 7 Making the Land 8 The First People 10 People from the Sea 12 James Morrill 15 Farmers 17 Taking the Land 20 A Port for Thuringowa 21 Travellers 23 Miners 25 The Great Northern Railway 28 Growth of a Community 30 Closer Settlement 32 Towns 34 Sugar 36 New Industries 39 Empires 43 We can be our country 45 Federation 46 War in Europe 48 Depression 51 War in the North 55 The Americans Arrive 57 Prosperous Times 63 A great city 65 Bibliography 69 Index 74 Photograph Index 78 gOrtvard To celebrate our nations Centenary, and the various Thuringowan communities' contribution to our sense of nation, this book was commissioned. Two previous council publications, Thuringowa Past and Present and It Was a Different Town have been modest, yet tantalising introductions to facets of our past. -

Doh Multi-Page Template
Medical Entomology 2016/2017 Annual Report Environmental Health Hazards Unit Environmental Health Directorate Public Health Division Department of Health, Western Australia PO Box 8172, Perth Business Centre Perth, 6849 Acknowledgements The extensive and diverse work program undertaken by the Department of Health’s Medical Entomology team outlined in this Annual Report could not have been completed without significant assistance and collaboration from many partners and stakeholders. In particular, the Medical Entomology team wishes to thank the Department of Health for its ongoing support in regards to this important public health program including: o the Environmental Health Directorate; o the Communicable Disease Control Directorate; o Population Health Units/Area Health Services o PathWest; and o Communications Directorate. The Pathwest laboratory continued to have a significant involvement in the program through provision of laboratory services for key components of the surveillance program for detection of arboviruses of public health significance to the State. We also acknowledge and thank the Population Health Units and the Western Australian Country Health Service for their role in reporting and follow-up of human cases of disease, and especially the role of Local Governments in the management of mosquitoes and the diseases they transmit. These organisations play an active role in the provision of data, case follow up investigations, care and bleeding of chickens for the sentinel chicken program, trapping of mosquitoes, mosquito control treatments and advice to the Western Australian community about disease risk through the media. In particular we thank Environmental Health Officers from the 139 Local Governments across WA, especially those within Contiguous Local Authority Groups (CLAGs), who respond to public complaints, undertake larval and adult mosquito surveys, and undertake mosquito control activities as part of their complex, integrated programs to manage the risks to public health and amenity within their regions. -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

WQ1181 - Ros! S River Basin
! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! Bohle River ! ! Stony Creek ! ! estuarine waters Town Common ! estuarine waters ! ! fresh waters ! ! ! ! ! Town Common ! ! ! estuarine waters ! ! ek ! e Cr ! ! ! r ! ! te a ! w ! ! ! *# e u ! l ek ! B re ! C ! ! ! ! ! ! ! ! y l a ! k e ! ! e r e Pallarenda fresh waters C H k ! ! ! ! ! e e ! ! r ! ! ! Deep C Cleveland Red Rock ! ! ! k ic l Pallarenda ! ! ! Bay A estuarine waters Ross Creek Bay ! ! Cape Cleveland ! estuarine waters ! ! fresh waters ! r ! ! e ! v ! ! i ! r R e ! v ! ! i R Rowes ! ! k c a e l l ! ! B Bay -
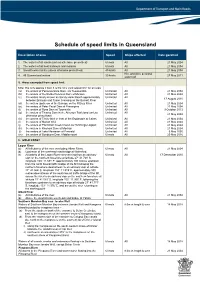
Schedule of Speed Limits in Queensland
Schedule of speed limits in Queensland Description of area Speed Ships affected Date gazetted 1. The waters of all canals (unless otherwise prescribed) 6 knots All 21 May 2004 2. The waters of all boat harbours and marinas 6 knots All 21 May 2004 3. Smooth water limits (unless otherwise prescribed) 40 knots All 21 May 2004 Hire and drive personal 4. All Queensland waters 30 knots 27 May 2011 watercraft 5. Areas exempted from speed limit Note: this only applies if item 3 is the only valid speed limit for an area (a) the waters of Perserverance Dam, via Toowoomba Unlimited All 21 May 2004 (b) the waters of the Bjelke Peterson Dam at Murgon Unlimited All 21 May 2004 (c) the waters locally known as Sandy Hook Reach approximately Unlimited All 17 August 2010 between Branyan and Tyson Crossing on the Burnett River (d) the waters upstream of the Barrage on the Fitzroy River Unlimited All 21 May 2004 (e) the waters of Peter Faust Dam at Proserpine Unlimited All 21 May 2004 (f) the waters of Ross Dam at Townsville Unlimited All 9 October 2013 (g) the waters of Tinaroo Dam in the Atherton Tableland (unless Unlimited All 21 May 2004 otherwise prescribed) (h) the waters of Trinity Inlet in front of the Esplanade at Cairns Unlimited All 21 May 2004 (i) the waters of Marian Weir Unlimited All 21 May 2004 (j) the waters of Plantation Creek known as Hutchings Lagoon Unlimited All 21 May 2004 (k) the waters in Kinchant Dam at Mackay Unlimited All 21 May 2004 (l) the waters of Lake Maraboon at Emerald Unlimited All 6 May 2005 (m) the waters of Bundoora Dam, Middlemount 6 knots All 20 May 2016 6. -

Status of Non-Native Freshwater Fishes in Tropical Northern
Journal & Proceedings of the Royal Society of New South Wales, Vol. 140, p. 63–78, 2007 ISSN 0035-9173/07/020063–16 $4.00/1 Status of Non-native Freshwater Fishes in Tropical Northern Queensland, Including Establishment Success, Rates of Spread, Range and Introduction Pathways alan charles webb Abstract: At least 20 non-native fishes have been reported from northern Queensland fresh waters, a 75% increase since 1994. Eleven of these species have established breeding populations and some are locally abundant and highly invasive, such as the tilapiine cichlids (Oreochromis mossambicus and Tilapia mariae) and the poeciliids (Gambusia holbrooki and Poecilia reticulata). Besides the continued introduction of non-native species, of great concern is the further spread of the tilapias, especially Oreochromis mossambicus and its hybrid form, and of another invasive, the three-spot gourami, Trichopterus trichogaster. Initial introductions are most probably releases of unwanted aquarium fish directly into open waters, or indirectly from ornamental ponds by flood waters. While natural dispersal is occurring, most of the range expansion of the tilapiine cichlids, particularly into impoundments in flood- prone areas, has been as a result of human translocation, and possibly the use of live bait by anglers. Keywords: Cichlidae, distribution patterns, Gambusia, Gourami, introduction pathways, invasive fishes, Oreochromis mossambicus, Poeciliidae, Tilapia INTRODUCTION native fishes in northern Queensland (McKay (1978, 1989, Arthington et al. 1984, Lear 1987), The history of non-native fishes, i.e., those orig- while McKay (1989) also referred to a previ- inating from overseas, introduced into northern ous, though unsuccessful, introduction of Jor- Queensland fresh waters has been well docu- danella sp. -

Mosquito-Borne Viruses and Non-Human Vertebrates in Australia: a Review
viruses Review Mosquito-Borne Viruses and Non-Human Vertebrates in Australia: A Review Oselyne T. W. Ong 1,2 , Eloise B. Skinner 3,4 , Brian J. Johnson 2 and Julie M. Old 5,* 1 Children’s Medical Research Institute, Westmead, NSW 2145, Australia; [email protected] 2 Mosquito Control Laboratory, QIMR Berghofer Medical Research Institute, Herston, QLD 4006, Australia; [email protected] 3 Environmental Futures Research Institute, Griffith University, Gold Coast, QLD 4222, Australia; [email protected] 4 Biology Department, Stanford University, Stanford, CA 94305, USA 5 School of Science, Western Sydney University, Hawkesbury, Locked bag 1797, Penrith, NSW 2751, Australia * Correspondence: [email protected] Abstract: Mosquito-borne viruses are well recognized as a global public health burden amongst humans, but the effects on non-human vertebrates is rarely reported. Australia, houses a number of endemic mosquito-borne viruses, such as Ross River virus, Barmah Forest virus, and Murray Valley encephalitis virus. In this review, we synthesize the current state of mosquito-borne viruses impacting non-human vertebrates in Australia, including diseases that could be introduced due to local mosquito distribution. Given the unique island biogeography of Australia and the endemism of vertebrate species (including macropods and monotremes), Australia is highly susceptible to foreign mosquito species becoming established, and mosquito-borne viruses becoming endemic alongside novel reservoirs. For each virus, we summarize the known geographic distribution, mosquito vectors, vertebrate hosts, clinical signs and treatments, and highlight the importance of including non-human vertebrates in the assessment of future disease outbreaks. The mosquito-borne viruses discussed can impact wildlife, livestock, and companion animals, causing significant changes to Australian Citation: Ong, O.T.W.; Skinner, E.B.; ecology and economy.