Cynoscion Arenarius and C.Nebulosus,The White
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Practical Handbook for Determining the Ages of Gulf of Mexico And
A Practical Handbook for Determining the Ages of Gulf of Mexico and Atlantic Coast Fishes THIRD EDITION GSMFC No. 300 NOVEMBER 2020 i Gulf States Marine Fisheries Commission Commissioners and Proxies ALABAMA Senator R.L. “Bret” Allain, II Chris Blankenship, Commissioner State Senator District 21 Alabama Department of Conservation Franklin, Louisiana and Natural Resources John Roussel Montgomery, Alabama Zachary, Louisiana Representative Chris Pringle Mobile, Alabama MISSISSIPPI Chris Nelson Joe Spraggins, Executive Director Bon Secour Fisheries, Inc. Mississippi Department of Marine Bon Secour, Alabama Resources Biloxi, Mississippi FLORIDA Read Hendon Eric Sutton, Executive Director USM/Gulf Coast Research Laboratory Florida Fish and Wildlife Ocean Springs, Mississippi Conservation Commission Tallahassee, Florida TEXAS Representative Jay Trumbull Carter Smith, Executive Director Tallahassee, Florida Texas Parks and Wildlife Department Austin, Texas LOUISIANA Doug Boyd Jack Montoucet, Secretary Boerne, Texas Louisiana Department of Wildlife and Fisheries Baton Rouge, Louisiana GSMFC Staff ASMFC Staff Mr. David M. Donaldson Mr. Bob Beal Executive Director Executive Director Mr. Steven J. VanderKooy Mr. Jeffrey Kipp IJF Program Coordinator Stock Assessment Scientist Ms. Debora McIntyre Dr. Kristen Anstead IJF Staff Assistant Fisheries Scientist ii A Practical Handbook for Determining the Ages of Gulf of Mexico and Atlantic Coast Fishes Third Edition Edited by Steve VanderKooy Jessica Carroll Scott Elzey Jessica Gilmore Jeffrey Kipp Gulf States Marine Fisheries Commission 2404 Government St Ocean Springs, MS 39564 and Atlantic States Marine Fisheries Commission 1050 N. Highland Street Suite 200 A-N Arlington, VA 22201 Publication Number 300 November 2020 A publication of the Gulf States Marine Fisheries Commission pursuant to National Oceanic and Atmospheric Administration Award Number NA15NMF4070076 and NA15NMF4720399. -

Andrea RAZ-GUZMÁN1*, Leticia HUIDOBRO2, and Virginia PADILLA3
ACTA ICHTHYOLOGICA ET PISCATORIA (2018) 48 (4): 341–362 DOI: 10.3750/AIEP/02451 AN UPDATED CHECKLIST AND CHARACTERISATION OF THE ICHTHYOFAUNA (ELASMOBRANCHII AND ACTINOPTERYGII) OF THE LAGUNA DE TAMIAHUA, VERACRUZ, MEXICO Andrea RAZ-GUZMÁN1*, Leticia HUIDOBRO2, and Virginia PADILLA3 1 Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Ciudad de México 2 Instituto Nacional de Pesca y Acuacultura, SAGARPA, Ciudad de México 3 Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad de México Raz-Guzmán A., Huidobro L., Padilla V. 2018. An updated checklist and characterisation of the ichthyofauna (Elasmobranchii and Actinopterygii) of the Laguna de Tamiahua, Veracruz, Mexico. Acta Ichthyol. Piscat. 48 (4): 341–362. Background. Laguna de Tamiahua is ecologically and economically important as a nursery area that favours the recruitment of species that sustain traditional fisheries. It has been studied previously, though not throughout its whole area, and considering the variety of habitats that sustain these fisheries, as well as an increase in population growth that impacts the system. The objectives of this study were to present an updated list of fish species, data on special status, new records, commercial importance, dominance, density, ecotic position, and the spatial and temporal distribution of species in the lagoon, together with a comparison of Tamiahua with 14 other Gulf of Mexico lagoons. Materials and methods. Fish were collected in August and December 1996 with a Renfro beam net and an otter trawl from different habitats throughout the lagoon. The species were identified, classified in relation to special status, new records, commercial importance, density, dominance, ecotic position, and spatial distribution patterns. -

02 Anderson FB107(1).Indd
Evolutionary associations between sand seatrout (Cynoscion arenarius) and silver seatrout (C. nothus) inferred from morphological characters, mitochondrial DNA, and microsatellite markers Item Type article Authors Anderson, Joel D.; McDonald, Dusty L.; Sutton, Glen R.; Karel, William J. Download date 23/09/2021 08:57:20 Link to Item http://hdl.handle.net/1834/25457 13 Abstract—The evolutionary asso- ciations between closely related fish Evolutionary associations between sand seatrout species, both contemporary and his- (Cynoscion arenarius) and silver seatrout torical, are frequently assessed by using molecular markers, such as (C. nothus) inferred from morphological characters, microsatellites. Here, the presence mitochondrial DNA, and microsatellite markers and variability of microsatellite loci in two closely related species of marine fishes, sand seatrout (Cynoscion are- Joel D. Anderson (contact author)1 narius) and silver seatrout (C. nothus), Dusty L. McDonald1 are explored by using heterologous primers from red drum (Sciaenops Glen R. Sutton2 ocellatus). Data from these loci are William J. Karel1 used in conjunction with morphologi- cal characters and mitochondrial DNA Email address for contact author: [email protected] haplotypes to explore the extent of 1 Perry R. Bass Marine Fisheries Research Station genetic exchange between species off- Texas Parks and Wildlife Department shore of Galveston Bay, TX. Despite HC02 Box 385, Palacios, Texas 77465 seasonal overlap in distribution, low 2 Galveston Bay Field Office genetic divergence at microsatellite Texas Parks and Wildlife Department loci, and similar life history param- 1502 FM 517 East, eters of C. arenarius and C. nothus, all Dickinson, Texas 77539 three data sets indicated that hybrid- ization between these species does not occur or occurs only rarely and that historical admixture in Galveston Bay after divergence between these species was unlikely. -

Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes
Old Dominion University ODU Digital Commons Biological Sciences Theses & Dissertations Biological Sciences Summer 2016 Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes Christi Linardich Old Dominion University, [email protected] Follow this and additional works at: https://digitalcommons.odu.edu/biology_etds Part of the Biodiversity Commons, Biology Commons, Environmental Health and Protection Commons, and the Marine Biology Commons Recommended Citation Linardich, Christi. "Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes" (2016). Master of Science (MS), Thesis, Biological Sciences, Old Dominion University, DOI: 10.25777/hydh-jp82 https://digitalcommons.odu.edu/biology_etds/13 This Thesis is brought to you for free and open access by the Biological Sciences at ODU Digital Commons. It has been accepted for inclusion in Biological Sciences Theses & Dissertations by an authorized administrator of ODU Digital Commons. For more information, please contact [email protected]. HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES by Christi Linardich B.A. December 2006, Florida Gulf Coast University A Thesis Submitted to the Faculty of Old Dominion University in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE BIOLOGY OLD DOMINION UNIVERSITY August 2016 Approved by: Kent E. Carpenter (Advisor) Beth Polidoro (Member) Holly Gaff (Member) ABSTRACT HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES Christi Linardich Old Dominion University, 2016 Advisor: Dr. Kent E. Carpenter Understanding the status of species is important for allocation of resources to redress biodiversity loss. -

Diets of Coastal Bottlenose Dolphins from the U. S. Mid
MARINE MAMMAL SCIENCE, 20(3):527-545 (July 2004) 0 2004 by the Society for Marine Mammalogy DIETS OF COASTAL BOTTLENOSE DOLPHINS FROM THE U.S. MID-ATLANTIC COAST DIFFER BY HABITAT DAMONP. GANNON' DANIELLEM. WAPLES Duke University Marine Laboratory, Nicholas School of the Environment and Earth Sciences, Duke University, 135 Duke Marine Lab Road, Beaufort, North Carolina 28516, U.S.A. E-mail: [email protected] ABSTRACT We recorded 31 species in the stomachs of 146 coastal bottlenose dolphins (Tursiops truncatu) from North Carolina, U.S.A. Sciaenid fishes were the most common prey (frequency of occurrence = 95%). By mass, Atlantic croaker (Micropogonias undulatus) dominated the diet of dolphins that stranded inside estuaries, whereas weakfish (Cynosicon regalis) was most important for dolphins in the ocean. Inshore squid (Loligo sp.) was eaten commonly by dolphins in the ocean, but not by those in the estuaries. There was no significant pattern in prey size associated with dolphin demography, but the proportion of the diet represented by croaker was higher for males than for females, and mature dolphins ate more croaker than did juveniles. Dietary differences between dolphins that stranded in the estuaries and those that stranded on ocean beaches support the hypothesis that some members of the population inhabit the ocean primarily while others reside principally in estuaries. The overwhelming majority of prey were soniferous species (75% of numerical abundance), which is consistent with the hypothesis that bottlenose dolphins use passive listening to locate noise-making fishes. However, spatiotemporal patterns in consumption of Sciaenid fishes did not coincide with their spawning, which is when peak sound production is thought to occur. -

The Sea Trout of Weakfishes of the Gulf of Mexico 1958
··· OFHCE COPY ONlY 11. I· I I I \ ,• . TECHNICAL SUMMAltY··N.9 .. l .. · .. / ) / ! -!·1 . I /"". THE"· SEA• :TR.OUT~ • •, • •6R • WEAKFISHES• • • • J ' I . -y~ , ... --:- ',1 j (GENUS· CYNOSCION)., ~- I ( --.- I~·' i ' '·r - _..,, 1 _.. i ' OF THE \GULF-'OF .ME-XICO . , I ' ••. / • ' .. '1 />' ~·:. / ,'\ ·. '~.::-'<·<· :.',, . '\ /; \ ! 1,. i'. .. , '•' ' 1 \ /' by • ; • I ' • '~ •• ' i". - ·:, ·I., .J WILLIAM :<Z. GUEST "Texas. Garn~ and 'Fish' comrtii~sim11 1 · Rockport, .Texas ···; anq ·,.... , ·, GbRDON1 G$TER. \ GulfCgast :R..eseatch Laboratory Ocea~ Sp.rings, Mis's1ssipp~ . '/ " OCTOBER, . 1-958 , I ( ! ·)' ; \ , I . 'J \ .. , j\ )'.,I i ·,1 . I /... The Gulf . .States Marine :Fisheries Commis- , ' .' ' ' -.._ \ . \ sion realizing that .data on the sp'.eckled trout I \ i and tlie,tw:o;:~pecies of white trout appearing ' i ' in waters of the Gulf states· should ·b~ sum- . , 1 ·marized, is pleasea to P,resent this · .publi~ / \' cation. · ·· . '1-. Data, appearing>' herein have been gathered: from a multipl.icity of sources, both published ·-- . and iunp:ubll~hetl, as is ·evidenced by the ac- . · companying ci,tation·s. It is believed the basic · information ~ontained in this publication ' \ · can be of considerable assistance to state marine fishery legisiatiye committees "and · . ' state fishery agencies in consideration of.. · ' I management measures designed to preserve i I :I these .s'p'ecies for the "COm:tnercial and sport fishermen of bot~ the present and tlie future. ' d J I ,I ir,-- I I I ) i, ' \ Ii I l ~ -Oiulf ~fates )mtarine j'Jiisheries illomtnission ·1?·. I ' 1·, 1 ,I. ,) TECHNICAL SUMMARY No. 1 I ) , I JI ~ ,) .. : .\ ' I 'THE SEA TROUT OR WEAKFISHES (GENUS CYNOSCION) OF THE GULF OF MEXICO by WILLIAM C. -

Living Resources Report Texas A&M University-Corpus Christi Results - Open Bay Habitat
Center for Coastal Studies CCBNEP Living Resources Report Texas A&M University-Corpus Christi Results - Open Bay Habitat B. Living Resources - Habitats Detailed community profiles of estuarine habitats within the CCBNEP study area are not available. Therefore, in the following sections, the organisms, community structure, and ecosystem processes and functions of the major estuarine habitats (Open Bay, Oyster Reef, Hard Substrate, Seagrass Meadow, Coastal Marsh, Tidal Flat, Barrier Island, and Gulf Beach) within the CCBNEP study area are presented. The following major subjects will be addressed for each habitat: (1) Physical setting and processes; (2) Producers and Decomposers; (3) Consumers; (4) Community structure and zonation; and (5) Ecosystem processes. HABITAT 1: OPEN BAY Table Of Contents Page 1.1. Physical Setting & Processes ............................................................................ 45 1.1.1 Distribution within Project Area ......................................................... 45 1.1.2 Historical Development ....................................................................... 45 1.1.3 Physiography ...................................................................................... 45 1.1.4 Geology and Soils ................................................................................ 46 1.1.5 Hydrology and Chemistry ................................................................... 47 1.1.5.1 Tides .................................................................................... 47 1.1.5.2 Freshwater -

Report T-668 Population Characteristics, Food Habits and Spawning Activity of Spotted Seatrout, Cvnoscion Nebulosus, in EVE R
Report T-668 Population Characteristics, Food Habits and Spawning Activity of Spotted Seatrout, Cvnoscion nebulosus, in EVE R NATIONAL -r., c, 3 01.... "C*, 0, @r*, lnI.10. Everglades National Park, South Flor~daResearch Center, P.O. Box 279, Homestead, Florida 33030 Population Characteristics, Food Habits and Spawning Activity of Spotted Seatrout, Cynoscion nebulosus, in Everglades National Park, Florida Report T-668 Edward Rutherford, Edith Thue and David Buker National Park Service South Florida Research Center Everglades National Park Homestead, Florida 33030 June 1982 TABLE OF CONTENTS LIST OF TABLES ........................... ii LIST OF FIGURES .......................... iii LIST OF APPENDICES ......................... v ABSTRACT ............................. 1 INTRODUCTION ........................... 2 Description of the Study Area ................... 3 METHODS .............................. 3 Data Collection ......................... 3 Aging Methods .......................... 3 Growth ............................. 4 Survival ............................ 5 Food Analyses .......................... 5 RESULTS .............................. 5 Length Frequency ........................ 6 Verification of Aging Technique .................. 6 Age Composition and Sex Ratio of the Catch ............ 7 Growth ............................. 8 Growth Equation ......................... 9 Length-weight Relationship .................... 9 Survival ............................. 9 Food Analyses .......................... 10 Spawning Activity ....................... -

Ontogenetic Variation in Sciaenid Otolith Morphometry with Fish Size from the Northern Gulf of Mexico
Nova Southeastern University NSUWorks All HCAS Student Capstones, Theses, and Dissertations HCAS Student Theses and Dissertations 1-7-2021 Ontogenetic Variation in Sciaenid Otolith Morphometry with Fish Size from the Northern Gulf of Mexico Thomas C. Ingalls Nova Southeastern University Follow this and additional works at: https://nsuworks.nova.edu/hcas_etd_all Part of the Aquaculture and Fisheries Commons, and the Marine Biology Commons Share Feedback About This Item NSUWorks Citation Thomas C. Ingalls. 2021. Ontogenetic Variation in Sciaenid Otolith Morphometry with Fish Size from the Northern Gulf of Mexico. Master's thesis. Nova Southeastern University. Retrieved from NSUWorks, . (34) https://nsuworks.nova.edu/hcas_etd_all/34. This Thesis is brought to you by the HCAS Student Theses and Dissertations at NSUWorks. It has been accepted for inclusion in All HCAS Student Capstones, Theses, and Dissertations by an authorized administrator of NSUWorks. For more information, please contact [email protected]. Thesis of Thomas C. Ingalls Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science Marine Science Nova Southeastern University Halmos College of Arts and Sciences January 2021 Approved: Thesis Committee Major Professor: Rosanna J. Milligan, Ph.D. Committee Member: Paul Arena, Ph.D. Committee Member: Philip Matich, Ph.D. This thesis is available at NSUWorks: https://nsuworks.nova.edu/hcas_etd_all/34 HALMOS COLLEGE OF ARTS AND SCIENCES Ontogenetic Variation in Sciaenid Otolith Morphometry with Fish Size from the Northern Gulf of Mexico By Thomas C. Ingalls Submitted to the Faculty of Halmos College of Arts and Sciences in partial fulfillment of the requirements for the degree of Master of Science with a specialty in: Marine Biology Nova Southeastern University 2021 1 Thesis of Thomas C. -

Download Vol. 56, No. 3
BULLETIN of the Florida Museum of Natural History TELEOSTEAN OTOLITHS REVEAL DIVERSE PLIO- PLEISTOCENE FISH ASSEMBLAGES IN COASTAL GEORGIA (GLYNN COUNTY) Gary L. Stringer and Dennis Bell Vol. 56, No. 3, pp. 83–108 August 9, 2018 ISSN 2373-9991 UNIVERSITY OF FLORIDA GAINESVILLE The FLORIDA MUSEUM OF NATURAL HISTORY is Florida’s state museum of natural history, dedicated to understanding, preserving, and interpreting biological diversity and cultural heritage. The BULLETIN OF THE FLORIDA MUSEUM OF NATURAL HISTORY is an on-line, open-ac- cess, peer-reviewed journal that publishes results of original research in zoology, botany, paleontology, archaeology, and museum science. New issues of the Bulletin are published at irregular intervals, and volumes are not necessarily completed in any one year. Volumes contain between 150 and 300 pages, sometimes more. The number of papers contained in each volume varies, depending upon the number of pages in each paper, but four numbers is the current standard. Multi-author issues of related papers have been published together, and inquiries about putting together such issues are welcomed. Address all inqui- ries to the Editor of the Bulletin. The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN (http://zoobank.org/). The ZooBank Publication number for this issue is EB7556D6-823A-470D-813F-8AC26650EC89. Richard C. Hulbert Jr., Editor Bulletin Committee Richard C. -

Aspects of the Biology of the Spotted Seatrout, Cynoscion Nebulosus, in Mississippi
Gulf and Caribbean Research Volume 7 Issue 5 Volume 7, Supplement 1 January 1983 Aspects of the Biology of the Spotted Seatrout, Cynoscion nebulosus, in Mississippi Robin M. Overstreet Gulf Coast Research Laboratory Follow this and additional works at: https://aquila.usm.edu/gcr Part of the Marine Biology Commons Recommended Citation Overstreet, R. M. 1983. Aspects of the Biology of the Spotted Seatrout, Cynoscion nebulosus, in Mississippi. Gulf Research Reports 7 (S1): 1-43. Retrieved from http://aquila.usm.edu/gcr/vol7/iss4/1 This Article is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf and Caribbean Research by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. GulfResearch Reports, Supplement 1, 1-43, June 1983 IC6 6 ASPECTS OF THE BIOLOGY OF THE SPOTTED SEATROUT, CYNOSCION NEBULOSUS, IN MISSISSIPPI ROBIN M. OVERSTREET GulfCoast Research Laboratory, Ocean Springs, Mississippi 39564 ABSTRACT About 3,000 specimens of the spotted seatrout from Mississippi Sound and adjacent water grouped by males and females had a nearly identical standard length (SL) versus total length (TL) relationship, although the equation for males in winter differed from that for those in other seasons. When investigating the SL-weight relationship, some differences occurred both among seasons and between sexes. Therefore, condition coefficients (K) were calculated to compare male and female groups according to their length and state of maturation on a seasonal basis. The hepatosomatic index (HSI) tended to increase with fish length, with relatively high values occurring in winter compared to low ones in summer when livers exhibited an abundance of lipid. -
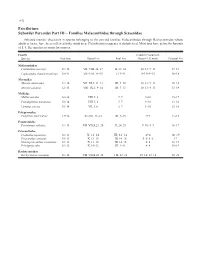
Perciformes Suborder Percoidei Part III – Families Malacanthidae
1152 Perciformes Suborder Percoidei Part III – Families Malacanthidae through Sciaenidae Selected meristic characters in species belonging to the percoid families Malacanthidae through Rachycentridae whose adults or larvae have been collected in the study area. Classification sequence is alphabetical. Most taxa have pelvic fin formula of I, 5. See species accounts for sources. Family Caudal (Procurrent, Species Vertebrae Dorsal Fin Anal Fin Dorsal + Ventral) Pectoral Fin Malacanthidae Caulolatilus microps 11+16 VII–VIII, 24–27 II, 22–24 10–13+9–13 17–18 Lopholatilus chamaeleonticeps 10+14 VII–VIII, 14–15 I, 13–14 9–13+9–13 16–18 Moronidae Morone americana 11+14 VII–XI, I, 11–13 III, 9–10 10–13+9–13 10–18 Morone saxatilis 12+13 VIII–IX, I, 9–14 III, 7–13 10–13+9–13 13–19 Mullidae Mullus auratus 10+14 VIII, I, 8 I, 7 9+10 15–17 Pseudupeneus maculatus 10+14 VIII, I, 8 I, 7 9+10 13–16 Upeneus parvus 10+14 VII, I, 8 I, 7 9+10 15–16 Polyprionidae Polyprion americanus 13+14 XI–XII, 11–12 III, 9–10 9+9 17–18 Pomatomidae Pomatomus saltatrix 11+15 VII–VIII,I,23–28 II, 24–29 9–10+8–9 16–17 Priacanthidae Cookeolus japonicus 10+13 X, 12–14 III, 12–14 4+4 18–19 Priacanthus arenatus 10+13 X, 13–15 III, 14–16 5–6+5–6 17 Heteropriacanthus cruentatus 10+13 X, 13–14 III, 14–15 4+4 18–19 Pristigenys alta 10+13 X, 10–12 III, 9–11 4+4 16–19 Rachycentridae Rachycentron canadum 11+14 VII–VIII,I,26–34 I–II, 22–28 15–16+12–14 20–21 Early Stages of Fishes in the Western North Atlantic Ocean 1153 Perciformes Suborder Percoidei Part III – Families Malacanthidae through Sciaenidae Selected meristic characters in species belonging to the percoid family Sciaenidae whose adults or larvae have been col- lected in the study area.