Spatial Structure of Communities on Dead Pen Shells (Atrina Rigida) in Sea Grass Beds
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Preliminary Guide to the Identification of the Early Life History Stages
NOAA Technical Memorandum NMFS-SEFSC-416 PRELIMINARY GUIDE TO TIm IDENTIFICATION OF TIm EARLY LlFE mSTORY STAGES OF BLENNIOID FISHES OF THE WBSTHRN CENTR.AL.ATLANTIC, FAUNAL LIST ANI) MERISTIC DATA FOR All KNOWN BLENNIOID SPECIES gy MARrIN R. CAVALLUZZI AND JOHN E. OLNEY U.S. DEPARTMENT OF COMMERCE National Oceanic and Atniospheric Administration National Marine Fisheries Service Southeast Fisheries Science Center 75 Virginia Beach Drive Miami. Florida 33149 December 1998 NOAA Teclmical Memorandum NMFS-SEFSC-416 PRELlMINARY GUIDE TO TIlE IDBNTIFlCA110N OF TIlE EARLY LIFE HISTORY STAGES OF BLBNNIOm FISHES OF TIm WBSTBRN CBN'l'R.At·A11..ANi'IC, FAUNAL LIST AND MERISllC DATA" -. FOR ALL KNOWN BLBNNIOID SPECJBS BY ~TIN R. CAVALLUZZI AND JOHN E. OLNEY u.s. DBPAR'I'MffiIT OF COMMERCB William M:Daley, Secretary NatioDal Oceanic and Atmospheric Administration D. JIjDlCS Baker, Under Secretary for OCeaJI.Sand Atmosphere National Marine Fisheries Service , Rolland A. Scbmitten, Assistant Administrator for Fisheries December 1998 This Technical Memorandum series is Used for documentation and timely cot:mD1Urlcationofpreliminazy results, interim reports, or similar special-purpose information. Although the memoranda are not subject to complete formal review, editoPal control, or de1Biled editing, they are expected to reflect smmd professional work. NOTICE .The National Mariiie Fisheries Service (NMFS) does not approve, recommend or endorse any proprietary product or material mentioned in this publication. No reference shati be made to NMFS or to this publication furi:rished by NMFS, in any advertising or salespromoiion which would imply that NMFS approves, recommends, or endorses any proprietary product or proprietary material mentioned herein or which has as its purpose any mtent to cause directly or indirectly the advertised product to be used or purchased because of this NMFS publication. -

Betnune-Cookman Coll., Daytona Unclas Dedch, Fla.) 31 P HC $4.75 CSCL 06C G3/04 35571
.1-1-CR-1374G9) A SlUDY Of LAGOCNAi AND W74-20718 E 3UAPIIE PaOCESSES AFD ARTIFICIAL HALITATS I) IHE AREA OF THE JOHN F. KE,EEDY (Betnune-Cookman Coll., Daytona Unclas dedch, Fla.) 31 p HC $4.75 CSCL 06C G3/04 35571 A STUDY OF LAGOONAL AND ESTUARINE PROCESSES AND ARTIFICIAL HABITATS IN THE AREA OF THE JOHN F. KENNEDY SPACE CENTER By Premsukh Poonai A first annual report on a project conducted by Bethune-Cookman College under a financial grant made by the National Aeronautics and Space Administration September 1972 - October 1973 Reproduced by NATIONAL TECHNICAL INFORMATION SERVICE US Dopartment of Commerce Springfield, VA. 22151 NOTICE THIS DOCUMENT HAS BEEN REPRODUCED FROM THE BEST COPY FURNISHED US BY THE SPONSORING AGENCY. ALTHOUGH IT IS RECOGNIZED THAT CER- TAIN PORTIONS ARE ILLEGIBLE, IT IS BEING RE- LEASED IN THE INTEREST OF MAKING AVAILABLE AS MUCH INFORMATION AS POSSIBLE. TABLE OF COMTENTS Page Abstract 1 Introduction 2-3 Materials and methods 4-8 Figures 1-5 9-13 Results and discussion 14-17 Conclusions 18 Tables 1-6 19-24 Acknowledgements 25 Appendices 1-2 26-27 Bibliography 28-29 ABSTRACT In order to study the influence of an artificial habitat of discarded automobile tires upon the biomass in and around it, three sites were selected in the Banana River, one of which will serve as a control and the other two as locations for small tire reefs. One of the reefs has been established and the other is on the point of being laid down. Measurements and correlation studies of the biomasses and the species indicate that the Blodynamics of the sites are appreciably the same in the three cases, that there are probably adequate populations at the lower trophic levels, that there are perhaps redused numbers of upper level carnivores and that it is likely that small artiftkial havens can contribute to an lucreaso in populations of certain species of gmefish. -

A Two Parameter Discrete Lindley Distribution
Revista Colombiana de Estadística January 2016, Volume 39, Issue 1, pp. 45 to 61 DOI: http://dx.doi.org/10.15446/rce.v39n1.55138 A Two Parameter Discrete Lindley Distribution Distribución Lindley de dos parámetros Tassaddaq Hussain1;a, Muhammad Aslam2;b, Munir Ahmad3;c 1Department of Statistics, Government Postgraduate College, Rawalakot, Pakistan 2Department of Statistics, Faculty of Sciences, King Abdulaziz University, Jeddah, Saudi Arabia 3National College of Business Administration and Economics, Lahore, Pakistan Abstract In this article we have proposed and discussed a two parameter discrete Lindley distribution. The derivation of this new model is based on a two step methodology i.e. mixing then discretizing, and can be viewed as a new generalization of geometric distribution. The proposed model has proved itself as the least loss of information model when applied to a number of data sets (in an over and under dispersed structure). The competing mod- els such as Poisson, Negative binomial, Generalized Poisson and discrete gamma distributions are the well known standard discrete distributions. Its Lifetime classification, kurtosis, skewness, ascending and descending factorial moments as well as its recurrence relations, negative moments, parameters estimation via maximum likelihood method, characterization and discretized bi-variate case are presented. Key words: Characterization, Discretized version, Estimation, Geometric distribution, Mean residual life, Mixture, Negative moments. Resumen En este artículo propusimos y discutimos la distribución Lindley de dos parámetros. La obtención de este Nuevo modelo está basada en una metodo- logía en dos etapas: mezclar y luego discretizar, y puede ser vista como una generalización de una distribución geométrica. El modelo propuesto de- mostró tener la menor pérdida de información al ser aplicado a un cierto número de bases de datos (con estructuras de supra y sobredispersión). -

Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes
Old Dominion University ODU Digital Commons Biological Sciences Theses & Dissertations Biological Sciences Summer 2016 Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes Christi Linardich Old Dominion University, [email protected] Follow this and additional works at: https://digitalcommons.odu.edu/biology_etds Part of the Biodiversity Commons, Biology Commons, Environmental Health and Protection Commons, and the Marine Biology Commons Recommended Citation Linardich, Christi. "Hotspots, Extinction Risk and Conservation Priorities of Greater Caribbean and Gulf of Mexico Marine Bony Shorefishes" (2016). Master of Science (MS), Thesis, Biological Sciences, Old Dominion University, DOI: 10.25777/hydh-jp82 https://digitalcommons.odu.edu/biology_etds/13 This Thesis is brought to you for free and open access by the Biological Sciences at ODU Digital Commons. It has been accepted for inclusion in Biological Sciences Theses & Dissertations by an authorized administrator of ODU Digital Commons. For more information, please contact [email protected]. HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES by Christi Linardich B.A. December 2006, Florida Gulf Coast University A Thesis Submitted to the Faculty of Old Dominion University in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE BIOLOGY OLD DOMINION UNIVERSITY August 2016 Approved by: Kent E. Carpenter (Advisor) Beth Polidoro (Member) Holly Gaff (Member) ABSTRACT HOTSPOTS, EXTINCTION RISK AND CONSERVATION PRIORITIES OF GREATER CARIBBEAN AND GULF OF MEXICO MARINE BONY SHOREFISHES Christi Linardich Old Dominion University, 2016 Advisor: Dr. Kent E. Carpenter Understanding the status of species is important for allocation of resources to redress biodiversity loss. -

Trophic Transfer and Habitat Use of Oyster Crassostrea Virginica Reefs in Southwest Florida, Identified by Stable Isotope Analysis
Vol. 462: 125–142, 2012 MARINE ECOLOGY PROGRESS SERIES Published August 21 doi: 10.3354/meps09824 Mar Ecol Prog Ser Trophic transfer and habitat use of oyster Crassostrea virginica reefs in southwest Florida, identified by stable isotope analysis Holly A. Abeels1,2, Ai Ning Loh1, Aswani K. Volety1,* 1Florida Gulf Coast University, Fort Myers, Florida 33965, USA 2Present address: University of Florida/Institute of Food and Agricultural Sciences, Brevard County Extension Service, Cocoa, Florida 32926, USA ABSTRACT: Oyster reefs have been identified as essential fish habitat for resident and transient species. Many organisms found on oyster reefs, including shrimp, crabs, and small fishes, find shelter and food on the reef and in turn provide food for transient species that frequent oyster reefs. The objective of this study was to determine trophic transfer on oyster reefs in a subtropical environment using stable isotope compositions. Water, sediment, particulate organic matter, vari- ous crustaceans, fishes, as well as oysters were collected at 2 sites in Estero Bay, Florida, during the wet and dry seasons, and processed for δ13C and δ15N stable isotope analyses. Differences in freshwater input (salinity) resulted in differences in carbon and nitrogen isotope ratios. Overall, fish and shrimp are secondary consumers, with crabs and oysters as primary consumers, and organic matter sources at the lowest trophic level. Results of the study further demonstrate that reef-resident organisms consume other organisms found on the reef and/or primary -

Descriptive Statistics
IMGD 2905 Descriptive Statistics Chapter 3 Summarizing Data Q: how to summarize numbers? • With lots of playtesting, there is a lot of data (good!) • But raw data is often just a pile of numbers • Rarely of interest, or even sensible Summarizing Data Measures of central tendency Groupwork 4 3 7 8 3 4 22 3 5 3 2 3 • Indicate central tendency with one number? • What are pros and cons of each? Measure of Central Tendency: Mean • Aka: “arithmetic mean” or “average” =AVERAGE(range) =AVERAGEIF() – averages if numbers meet certain condition Measure of Central Tendency: Median • Sort values low to high and take middle value =MEDIAN(range) Measure of Central Tendency: Mode • Number which occurs most frequently • Not too useful in many cases Best use for categorical data • e.g., most popular Hero group in http://pad3.whstatic.com/images/thumb/c/cd/Find-the-Mode-of-a-Set-of-Numbers- Heroes of theStep-7.jpg/aid130521-v4-728px-Find-the-Mode-of-a-Set-of-Numbers-Step-7.jpg Storm =MODE() Mean, Median, Mode? frequency Mean, Median, Mode? frequency Mean, Median, Mode? frequency Mean, Median, Mode? frequency Mean, Median, Mode? frequency Mean, Median, Mode? mean median modes mode mean median frequency no mode mean (a) median (b) frequency mode mode median median (c) mean mean frequency frequency frequency (d) (e) Which to Use: Mean, Median, Mode? Which to Use: Mean, Median, Mode? • Mean many statistical tests that use sample ‒ Estimator of population mean ‒ Uses all data Which to Use: Mean, Median, Mode? • Median is useful for skewed data ‒ e.g., income data (US Census) or housing prices (Zillo) ‒ e.g., Overwatch team (6 players): 5 people level 5, 1 person level 275 ₊ Mean is 50 - not so useful since no one at this level ₊ Median is 5 – perhaps more representative ‒ Does not use all data. -

Salinity Tolerances for the Major Biotic Components Within the Anclote River and Anchorage and Nearby Coastal Waters
Salinity Tolerances for the Major Biotic Components within the Anclote River and Anchorage and Nearby Coastal Waters October 2003 Prepared for: Tampa Bay Water 2535 Landmark Drive, Suite 211 Clearwater, Florida 33761 Prepared by: Janicki Environmental, Inc. 1155 Eden Isle Dr. N.E. St. Petersburg, Florida 33704 For Information Regarding this Document Please Contact Tampa Bay Water - 2535 Landmark Drive - Clearwater, Florida Anclote Salinity Tolerances October 2003 FOREWORD This report was completed under a subcontract to PB Water and funded by Tampa Bay Water. i Anclote Salinity Tolerances October 2003 ACKNOWLEDGEMENTS The comments and direction of Mike Coates, Tampa Bay Water, and Donna Hoke, PB Water, were vital to the completion of this effort. The authors would like to acknowledge the following persons who contributed to this work: Anthony J. Janicki, Raymond Pribble, and Heidi L. Crevison, Janicki Environmental, Inc. ii Anclote Salinity Tolerances October 2003 EXECUTIVE SUMMARY Seawater desalination plays a major role in Tampa Bay Water’s Master Water Plan. At this time, two seawater desalination plants are envisioned. One is currently in operation producing up to 25 MGD near Big Bend on Tampa Bay. A second plant is conceptualized near the mouth of the Anclote River in Pasco County, with a 9 to 25 MGD capacity, and is currently in the design phase. The Tampa Bay Water desalination plant at Big Bend on Tampa Bay utilizes a reverse osmosis process to remove salt from seawater, yielding drinking water. That same process is under consideration for the facilities Tampa Bay Water has under design near the Anclote River. -

Observer Training Manual National Marine Fisheries Service Southeast
Characterization of the US Gulf of Mexico and Southeastern Atlantic Otter Trawl and Bottom Reef Fish Fisheries Observer Training Manual National Marine Fisheries Service Southeast Fisheries Science Center Galveston Laboratory September 2010 TABLE OF CONTENTS National Overview ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 1 Project Overview ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 8 Observer Program Guidelines and Safety ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 15 Observer Safety ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 15 Medical Fitness for Sea ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 15 Training ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 15 Before Deployment on Vessel ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 16 Seven Steps to Survival ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 18 Donning an Immersion Suit ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 20 Safety Aboard Vessels ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 22 Safety At‐Sea Transfers ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 23 Off‐Shore Communications ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 24 Advise to Women Going to Sea ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 27 Summary: What You Need to Know About Sea Survival ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 29 Deployment on Vessel -

The Need to Measure Variance in Experiential Learning and a New Statistic to Do So
Developments in Business Simulation & Experiential Learning, Volume 26, 1999 THE NEED TO MEASURE VARIANCE IN EXPERIENTIAL LEARNING AND A NEW STATISTIC TO DO SO John R. Dickinson, University of Windsor James W. Gentry, University of Nebraska-Lincoln ABSTRACT concern should be given to the learning process rather than performance outcomes. The measurement of variation of qualitative or categorical variables serves a variety of essential We contend that insight into process can be gained purposes in simulation gaming: de- via examination of within-student and across-student scribing/summarizing participants’ decisions for variance in decision-making. For example, if the evaluation and feedback, describing/summarizing role of an experiential exercise is to foster trial-and- simulation results toward the same ends, and error learning, the variance in a student’s decisions explaining variation for basic research purposes. will be greatest when the trial-and-error stage is at Extant measures of dispersion are not well known. its peak. To the extent that the exercise works as Too, simulation gaming situations are peculiar in intended, greater variances would be expected early that the total number of observations available-- in the game play as opposed to later, barring the usually the number of decision periods or the game world encountering a discontinuity. number of participants--is often small. Each extant measure of dispersion has its own limitations. On the other hand, it is possible that decisions may Common limitations are that, under ordinary be more consistent early and then more variable later circumstances and especially with small numbers if the student is trying to recover from early of observations, a measure may not be defined and missteps. -
Standard Deviation - Wikipedia Visited on 9/25/2018
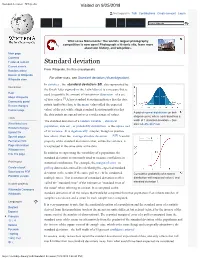
Standard deviation - Wikipedia Visited on 9/25/2018 Not logged in Talk Contributions Create account Log in Article Talk Read Edit View history Wiki Loves Monuments: The world's largest photography competition is now open! Photograph a historic site, learn more about our history, and win prizes. Main page Contents Featured content Standard deviation Current events Random article From Wikipedia, the free encyclopedia Donate to Wikipedia For other uses, see Standard deviation (disambiguation). Wikipedia store In statistics , the standard deviation (SD, also represented by Interaction the Greek letter sigma σ or the Latin letter s) is a measure that is Help used to quantify the amount of variation or dispersion of a set About Wikipedia of data values.[1] A low standard deviation indicates that the data Community portal Recent changes points tend to be close to the mean (also called the expected Contact page value) of the set, while a high standard deviation indicates that A plot of normal distribution (or bell- the data points are spread out over a wider range of values. shaped curve) where each band has a Tools The standard deviation of a random variable , statistical width of 1 standard deviation – See What links here also: 68–95–99.7 rule population, data set , or probability distribution is the square root Related changes Upload file of its variance . It is algebraically simpler, though in practice [2][3] Special pages less robust , than the average absolute deviation . A useful Permanent link property of the standard deviation is that, unlike the variance, it Page information is expressed in the same units as the data. -

Population Dispersion
AccessScience from McGraw-Hill Education Page 1 of 7 www.accessscience.com Population dispersion Contributed by: Francis C. Evans Publication year: 2014 The spatial distribution at any particular moment of the individuals of a species of plant or animal. Under natural conditions organisms are distributed either by active movements, or migrations, or by passive transport by wind, water, or other organisms. The act or process of dissemination is usually termed dispersal, while the resulting pattern of distribution is best referred to as dispersion. Dispersion is a basic characteristic of populations, controlling various features of their structure and organization. It determines population density, that is, the number of individuals per unit of area, or volume, and its reciprocal relationship, mean area, or the average area per individual. It also determines the frequency, or chance of encountering one or more individuals of the population in a particular sample unit of area, or volume. The ecologist therefore studies not only the fluctuations in numbers of individuals in a population but also the changes in their distribution in space. See also: POPULATION DISPERSAL. Principal types of dispersion The dispersion pattern of individuals in a population may conform to any one of several broad types, such as random, uniform, or contagious (clumped). Any pattern is relative to the space being examined; a population may appear clumped when a large area is considered, but may prove to be distributed at random with respect to a much smaller area. Random or haphazard. This implies that the individuals have been distributed by chance. In such a distribution, the probability of finding an individual at any point in the area is the same for all points (Fig. -

Literature EVALUATING GEAR and FACTORS AFFECTING CATCH
Literature EVALUATING GEAR AND FACTORS AFFECTING CATCH AND SAMPLING VARIATION Frank J. Schwartz Patricia A. Howland Special Scientific Report for .Carolina Power and Light Company Raleigh, North Carolina 1 June 1978 Institute of . Marine Sciences University of North Carolina Morehead City, North Carolina 28557 INTRODUCTION Scientists are always concerned with the data they obtain. This may be in the form of: did I sample enough, was the sample size large enough to be con- sidered representative, did I sample all habitats, faunas, etc., did I take the proper environmental data, and should I sample only during the day or night hours. These and a host of questions besiege him in his quest for representa- tive and precise information about the area or fauna being studied. These concerns are often unknowingly upset by ignoring factors other than biological that may seriously alter the results and conclusions reached. These may be subtle factors such as: how efficient is the gear used, how ef- fective is its operation, is it selective for certain sizes or species of organisms because of color or strength of material. While the organisms may behave one way in relation to the gear, how does the behavior of the gear affect the po- tentially capturable organism? What type of gear should be used--midwater or other? Will mesh size affect the results? Does one type of gear sample a species, population or area better than another and why? What effect does human fatigue have on gear operation? Even with awareness of these and many other factors that influence the outcome of a research study, extraneous factors beyond our control may fur- ther influence the observed data.