Tutorials and Reviews NEURAL EXCITABILITY, SPIKING AND
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Interactions Among Diameter, Myelination, and the Na/K Pump Affect Axonal Resilience to High-Frequency Spiking
Interactions among diameter, myelination, and the Na/K pump affect axonal resilience to high-frequency spiking Yunliang Zanga,b and Eve Mardera,b,1 aVolen National Center for Complex Systems, Brandeis University, Waltham, MA 02454; and bDepartment of Biology, Brandeis University, Waltham, MA 02454 Contributed by Eve Marder, June 22, 2021 (sent for review March 25, 2021; reviewed by Carmen C. Canavier and Steven A. Prescott) Axons reliably conduct action potentials between neurons and/or (16–21). It remains unknown how these directly correlated pro- other targets. Axons have widely variable diameters and can be cesses, fast spiking and slow Na+ removal, interact in the control myelinated or unmyelinated. Although the effect of these factors of reliable spike generation and propagation. Furthermore, it is on propagation speed is well studied, how they constrain axonal unclear whether reducing Na+-andK+-current overlap can con- resilience to high-frequency spiking is incompletely understood. sistently decrease the rate of Na+ influx and accordingly enhance Maximal firing frequencies range from ∼1Hzto>300 Hz across axonal reliability to propagate high-frequency spikes. Conse- neurons, but the process by which Na/K pumps counteract Na+ quently, by modeling the process of Na+ removal in unmyelinated influx is slow, and the extent to which slow Na+ removal is com- and myelinated axons, we systematically explored the effects of patible with high-frequency spiking is unclear. Modeling the pro- diameter, myelination, and Na/K pump density on spike propagation cess of Na+ removal shows that large-diameter axons are more reliability. resilient to high-frequency spikes than are small-diameter axons, because of their slow Na+ accumulation. -

Dynamics Forced by Homoclinic Orbits
DYNAMICS FORCED BY HOMOCLINIC ORBITS VALENT´IN MENDOZA Abstract. The complexity of a dynamical system exhibiting a homoclinic orbit is given by the orbits that it forces. In this work we present a method, based in pruning theory, to determine the dynamical core of a homoclinic orbit of a Smale diffeomorphism on the 2-disk. Due to Cantwell and Conlon, this set is uniquely determined in the isotopy class of the orbit, up a topological conjugacy, so it contains the dynamics forced by the homoclinic orbit. Moreover we apply the method for finding the orbits forced by certain infinite families of homoclinic horseshoe orbits and propose its generalization to an arbitrary Smale map. 1. Introduction Since the Poincar´e'sdiscovery of homoclinic orbits, it is known that dynamical systems with one of these orbits have a very complex behaviour. Such a feature was explained by Smale in terms of his celebrated horseshoe map [40]; more precisely, if a surface diffeomorphism f has a homoclinic point then, there exists an invariant set Λ where a power of f is conjugated to the shift σ defined on the compact space Σ2 = f0; 1gZ of symbol sequences. To understand how complex is a diffeomorphism having a periodic or homoclinic orbit, we need the notion of forcing. First we will define it for periodic orbits. 1.1. Braid types of periodic orbits. Let P and Q be two periodic orbits of homeomorphisms f and g of the closed disk D2, respectively. We say that (P; f) and (Q; g) are equivalent if there is an orientation-preserving homeomorphism h : D2 ! D2 with h(P ) = Q such that f is isotopic to h−1 ◦ g ◦ h relative to P . -

Energy Expenditure Computation of a Single Bursting Neuron
Cognitive Neurodynamics (2019) 13:75–87 https://doi.org/10.1007/s11571-018-9503-3 (0123456789().,-volV)(0123456789().,-volV) ORIGINAL ARTICLE Energy expenditure computation of a single bursting neuron 2 1,2 2 2 Fengyun Zhu • Rubin Wang • Xiaochuan Pan • Zhenyu Zhu Received: 17 October 2017 / Revised: 22 August 2018 / Accepted: 28 August 2018 / Published online: 3 September 2018 Ó The Author(s) 2018 Abstract Brief bursts of high-frequency spikes are a common firing pattern of neurons. The cellular mechanisms of bursting and its biological significance remain a matter of debate. Focusing on the energy aspect, this paper proposes a neural energy calculation method based on the Chay model of bursting. The flow of ions across the membrane of the bursting neuron with or without current stimulation and its power which contributes to the change of the transmembrane electrical potential energy are analyzed here in detail. We find that during the depolarization of spikes in bursting this power becomes negative, which was also discovered in previous research with another energy model. We also find that the neuron’s energy consumption during bursting is minimal. Especially in the spontaneous state without stimulation, the total energy con- sumption (2.152 9 10-7 J) during 30 s of bursting is very similar to the biological energy consumption (2.468 9 10-7 J) during the generation of a single action potential, as shown in Wang et al. (Neural Plast 2017, 2017a). Our results suggest that this property of low energy consumption could simply be the consequence of the biophysics of generating bursts, which is consistent with the principle of energy minimization. -

An Example of Bifurcation to Homoclinic Orbits
JOURNAL OF DIFFERENTIAL EQUATIONS 37, 351-373 (1980) An Example of Bifurcation to Homoclinic Orbits SHUI-NEE CHOW* Department of Mathematics, Michigan State University, East Lansing, Michigan 48823 AND JACK K. HALE+ AND JOHN MALLET-PARET Lefschetz Center for Dynamical Systems, Division of Applied Mathematics, Brown University, Providence, Rhode Island 02912 Received October 19, 1979; revised January 23, 1980 Consider the equation f - x + x 2 = -A12 + &f(t) where f(t + 1) = f(t) and h = (h, , A,) is small. For h = 0, there is a homoclinic orbit r through zero. For X # 0 and small, there can be “strange” attractors near r. The pur- pose of this paper is to determine the curves in /\-space of bifurcation to “strange” attractors and to relate this to hyperbolic subharmonic bifurcations. I. INTRODUCTION At the present time, there is considerable interest in the theory of “strange” attractors of mappings and the manner in which strange attractors arise through successive bifurcations of periodic orbits (see [l, 5-121). The simplest “strange” attractors arise near homoclinic points. For a hyperbolic fixed point p of a diffeomorphism, a point 4 # p is a homo- clinic point of p if 4 is in the intersection of the stable and unstable manifolds of p. The point 4 is called a transverse homoclinic point of p if the intersection of the stable and unstable manifolds of p at 4 is transversal. It was known to Poincare that the existence of one transverse homoclinic point implies there must be infinitely many transverse homoclinic points. Birkhoff showed that every * This research was supported in part by the Netional Science Foundation, under MCS76-06739. -

Characteristics of Hippocampal Primed Burst Potentiation in Vitro and in the Awake Rat
The Journal of Neuroscience, November 1988, 8(11): 4079-4088 Characteristics of Hippocampal Primed Burst Potentiation in vitro and in the Awake Rat D. M. Diamond, T. V. Dunwiddie, and G. M. Rose Medical Research Service, Veterans Administration Medical Center, and Department of Pharmacology, University of Colorado Health Sciences Center, Denver, Colorado 80220 A pattern of electrical stimulation based on 2 prominent (McNaughton, 1983; Lynch and Baudry, 1984; Farley and Al- physiological features of the hippocampus, complex spike kon, 1985; Byrne, 1987). Of these, a phenomenontermed long- discharge and theta rhythm, was used to induce lasting in- term potentiation (LTP) has received extensive attention be- creases in responses recorded in area CA1 of hippocampal causeit sharesa number of characteristicswith memory. For slices maintained in vitro and from the hippocampus of be- example, LTP has a rapid onset and long duration and is having rats. This effect, termed primed burst (PB) potentia- strengthenedby repetition (Barnes, 1979; Teyler and Discenna, tion, was elicited by as few as 3 stimuli delivered to the 1987). In addition, the decay of LTP is correlated with the time commissural/associational afferents to CAl. The patterns courseof behaviorally assessedforgetting (Barnes, 1979; Barnes of stimulus presentation consisted of a single priming pulse and McNaughton, 1985). The similarity of LTP and memory followed either 140 or 170 msec later by a high-frequency has fostered an extensive characterization of the conditions un- burst of 2-10 pulses; control stimulation composed of un- derlying the initiation of LTP, as well as its biochemical sub- primed high-frequency trains of up to 10 pulses had no en- strates (Lynch and Baudry, 1984; Wigstrijm and Gustafsson, during effect. -

The Logic Behind Neural Control of Breathing Pattern Alona Ben-Tal 1, Yunjiao Wang2 & Maria C
www.nature.com/scientificreports OPEN The logic behind neural control of breathing pattern Alona Ben-Tal 1, Yunjiao Wang2 & Maria C. A. Leite3 The respiratory rhythm generator is spectacular in its ability to support a wide range of activities and Received: 10 January 2019 adapt to changing environmental conditions, yet its operating mechanisms remain elusive. We show Accepted: 29 May 2019 how selective control of inspiration and expiration times can be achieved in a new representation of Published: xx xx xxxx the neural system (called a Boolean network). The new framework enables us to predict the behavior of neural networks based on properties of neurons, not their values. Hence, it reveals the logic behind the neural mechanisms that control the breathing pattern. Our network mimics many features seen in the respiratory network such as the transition from a 3-phase to 2-phase to 1-phase rhythm, providing novel insights and new testable predictions. Te mechanism for generating and controlling the breathing pattern by the respiratory neural circuit has been debated for some time1–6. In 1991, an area of the brainstem, the pre Bötzinger Complex (preBötC), was found essential for breathing7. An isolated single PreBötC neuron could generate tonic spiking (a non-interrupted sequence of action potentials), bursting (a repeating pattern that consists of a sequence of action potentials fol- lowed by a time interval with no action potentials) or silence (no action potentials)8. Tese signals are transmit- ted, through other populations of neurons, to spinal motor neurons that activate the respiratory muscle4,6. Te respiratory muscle contracts when it receives a sequence of action potentials from the motor neurons and relaxes when no action potentials arrive9. -

Attractors and Orbit-Flip Homoclinic Orbits for Star Flows
PROCEEDINGS OF THE AMERICAN MATHEMATICAL SOCIETY Volume 141, Number 8, August 2013, Pages 2783–2791 S 0002-9939(2013)11535-2 Article electronically published on April 12, 2013 ATTRACTORS AND ORBIT-FLIP HOMOCLINIC ORBITS FOR STAR FLOWS C. A. MORALES (Communicated by Bryna Kra) Abstract. We study star flows on closed 3-manifolds and prove that they either have a finite number of attractors or can be C1 approximated by vector fields with orbit-flip homoclinic orbits. 1. Introduction The notion of attractor deserves a fundamental place in the modern theory of dynamical systems. This assertion, supported by the nowadays classical theory of turbulence [27], is enlightened by the recent Palis conjecture [24] about the abun- dance of dynamical systems with finitely many attractors absorbing most positive trajectories. If confirmed, such a conjecture means the understanding of a great part of dynamical systems in the sense of their long-term behaviour. Here we attack a problem which goes straight to the Palis conjecture: The fini- tude of the number of attractors for a given dynamical system. Such a problem has been solved positively under certain circunstances. For instance, we have the work by Lopes [16], who, based upon early works by Ma˜n´e [18] and extending pre- vious ones by Liao [15] and Pliss [25], studied the structure of the C1 structural stable diffeomorphisms and proved the finitude of attractors for such diffeomor- phisms. His work was largely extended by Ma˜n´e himself in the celebrated solution of the C1 stability conjecture [17]. On the other hand, the Japanesse researchers S. -
![Codimension-Two Homoclinic Bifurcations Underlying Spike Adding in the Hindmarsh-Rose Burster Arxiv:1109.5689V1 [Math.DS] 26 S](https://docslib.b-cdn.net/cover/3701/codimension-two-homoclinic-bifurcations-underlying-spike-adding-in-the-hindmarsh-rose-burster-arxiv-1109-5689v1-math-ds-26-s-643701.webp)
Codimension-Two Homoclinic Bifurcations Underlying Spike Adding in the Hindmarsh-Rose Burster Arxiv:1109.5689V1 [Math.DS] 26 S
Codimension-two homoclinic bifurcations underlying spike adding in the Hindmarsh-Rose burster Daniele Linaro1, Alan Champneys2, Mathieu Desroches2, Marco Storace1 1Biophysical and Electronic Engineering Department, University of Genoa, Genova, Italy 2Department of Engineering Mathematics, University of Bristol, Bristol, UK July 2, 2018 Abstract The well-studied Hindmarsh-Rose model of neural action potential is revisited from the point of view of global bifurcation analysis. This slow-fast system of three paremeterised differential equations is arguably the simplest reduction of Hodgkin- Huxley models capable of exhibiting all qualitatively important distinct kinds of spiking and bursting behaviour. First, keeping the singular perturbation parameter fixed, a comprehensive two-parameter bifurcation diagram is computed by brute force. Of particular concern is the parameter regime where lobe-shaped regions of irregular bursting undergo a transition to stripe-shaped regions of periodic bursting. The boundary of each stripe represents a fold bifurcation that causes a smooth spike-adding transition where the number of spikes in each burst is increased by one. Next, numerical continuation studies reveal that the global structure is organised by various curves of homoclinic bifurcations. In particular the lobe to stripe transition is organised by a sequence of codimension- two orbit- and inclination-flip points that occur along each homoclinic branch. Each branch undergoes a sharp turning point and hence approximately has a double- cover of the same curve in parameter space. The sharp turn is explained in terms of the interaction between a two-dimensional unstable manifold and a one-dimensional slow manifold in the singular limit. Finally, a new local analysis is undertaken us- ing approximate Poincar´emaps to show that the turning point on each homoclinic branch in turn induces an inclination flip that gives birth to the fold curve that or- ganises the spike-adding transition. -

Homoclinic and Heteroclinic Bifurcations in Vector Fields
Homoclinic and Heteroclinic Bifurcations in Vector Fields Ale Jan Homburg Bj¨ornSandstede Korteweg{de Vries Institute for Mathematics Division of Applied Mathematics University of Amsterdam Brown University 1018 TV Amsterdam, The Netherlands Providence, RI 02906, USA June 17, 2010 Abstract An overview of homoclinic and heteroclinic bifurcation theory for autonomous vector fields is given. Specifically, homoclinic and heteroclinic bifurcations of codimension one and two in generic, equivariant, reversible, and conservative systems are reviewed, and results pertaining to the existence of multi-round homoclinic and periodic orbits and of complicated dynamics such as suspended horseshoes and attractors are stated. Bifurcations of homoclinic orbits from equilibria in local bifurcations are also considered. The main analytic and geometric techniques such as Lin's method, Shil'nikov variables and homoclinic center manifolds for analyzing these bifurcations are discussed. Finally, a few related topics, such as topological moduli, numerical algorithms, variational methods, and extensions to singularly perturbed and infinite- dimensional systems, are reviewed briefly. 1 Contents 1 Introduction 4 2 Homoclinic and heteroclinic orbits, and their geometry5 2.1 Homoclinic orbits to hyperbolic equilibria.............................6 2.2 Homoclinic orbits to nonhyperbolic equilibria........................... 10 2.3 Heteroclinic cycles with hyperbolic equilibria........................... 12 3 Analytical and geometric approaches 13 3.1 Normal forms and linearizability................................. -

Homoclinic Tangles for Noninvertible Maps
Nonlinear Analysis 41 (2000) 259–276 www.elsevier.nl/locate/na Homoclinic tangles for noninvertible maps Evelyn Sander Department of Mathematical Sciences, George Mason University, MS 3F2, 4400 University Drive, Fairfax, VA 22030, USA Received 2 March 1998; accepted 17 July 1998 Keywords: Homoclinic tangles; Noninvertible maps; Transverse homoclinic point 1. Introduction Not all naturally occurring iterated systems have the property that time is reversible. In which case, the dynamics must be modelled by a noninvertible map rather than by a di eomorphism. While one-dimensional theory describes dynamics of noninvertible maps, higher-dimensional theory has focussed on di eomorphisms, though there are many higher-dimensional examples of natural systems modelled by noninvertible maps. For example, noninvertible maps occur in the study of population dynamics [2], time one maps of delay equations [25], control theory algorithms [1, 5, 6], neural networks [16], and iterated di erence methods [14]. In order to use dynamical systems theory in applications for which maps are noninvertible, it is important to know how the theory di ers from the di eomorphism case. One of the principal indications of chaotic dynamics in deterministic systems is the existence of transverse homoclinic orbits. This paper examines the existence of homoclinic tangles and the associated dynamic behavior for noninvertible maps. In particular, we give examples of maps which have transverse intersection of stable and unstable manifolds but no horseshoe. Furthermore, we show that this is a codimension two phenomenon. Papers by Steinlein and Walther [24,25] and Lerman and Shil’nikov [13] both estab- lish conditions for a transverse homoclinic point of a noninvertible map to be contained in a horseshoe. -

Homoclinic Orbits in Hamiltonian Systems
CORE Metadata, citation and similar papers at core.ac.uk Provided by Elsevier - Publisher Connector JOURNAL OF DIFFERENTIAL EQUATIONS 21,431-438 (1976) Homoclinic Orbits in Hamiltonian Systems ROBERTL. DEVANEY Department of Mathematics, Northwestern University, Evanston, Illinois 60201 Received January 25, 1975 The object of this paper is to study the orbit structure of a Hamiltonian system in a neighborhood of a trajectory which is doubly asymptotic to an equilibrium solution, i.e., an orbit which lies in the intersection of the stable and unstable manifolds of a critical point. Such an orbit is called a homoclinic orbit. For diffeomorphisms, the analogous situation is fairly well understood. By a theorem of Smale [7], in every neighborhood of a transversal homoclinic point of a periodic point, there is a compact invariant set on which some iterate of the diffeomorphism is topologically conjugate to the Bernoulli shift on N symbols. Via Poincare maps on local transversal sections, there is thus an analogous result for hyperbolic closed orbits of vector fields. For critical points of vector fields, however, the situation is somewhat different. In the first place, by the Kupka-Smale theorem [4], the existence of orbits doubly asymptotic to an equilibrium point is not generic. In fact, the set of vector fields which admit homoclinic orbits at critical points is of the first Baire category in the set of all smooth vector fields. This follows since the sum of the dimensions of the stable and unstable manifolds at the critical point is at most equal to the dimension of the mani- fold itself. -
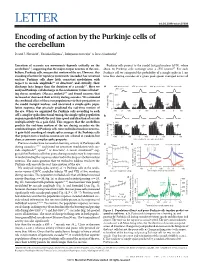
(2015) Encoding of Action by the Purkinje Cells of the Cerebellum
LETTER doi:10.1038/nature15693 Encoding of action by the Purkinje cells of the cerebellum David J. Herzfeld1, Yoshiko Kojima2, Robijanto Soetedjo2 & Reza Shadmehr1 Execution of accurate eye movements depends critically on the Purkinje cells project to the caudal fastigial nucleus (cFN), where cerebellum1–3, suggesting that the major output neurons of the cere- about 50 Purkinje cells converge onto a cFN neuron17. For each bellum, Purkinje cells, may predict motion of the eye. However, this Purkinje cell we computed the probability of a simple spike in 1-ms encoding of action for rapid eye movements (saccades) has remained time bins during saccades of a given peak speed, averaged across all unclear: Purkinje cells show little consistent modulation with respect to saccade amplitude4,5 or direction4, and critically, their discharge lasts longer than the duration of a saccade6,7.Herewe a 400° per s saccade 650° per s saccade 400° per s saccade 650° per s saccade analysed Purkinje-cell discharge in the oculomotor vermis of behav- 500° per s 8,9 200 ing rhesus monkeys (Macaca mulatta) and found neurons that N12 increased or decreased their activity during saccades. We estimated the combined effect of these two populations via their projections to 100 the caudal fastigial nucleus, and uncovered a simple-spike popu- lation response that precisely predicted the real-time motion of 0 the eye. When we organized the Purkinje cells according to each –1000 100 0 100 0 100 0 100 cell’s complex-spike directional tuning, the simple-spike population b 500° per s response predicted both the real-time speed and direction of saccade multiplicatively via a gain field.