High-Resolution Simulations of the Final Assembly of Earth-Like Planets 2: Water Delivery and Planetary Habitability
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Using Kepler Systems to Constrain the Frequency and Severity of Dynamical Effects on Habitable Planets Alexander James Mustill Melvyn B
Using Kepler systems to constrain the frequency and severity of dynamical effects on habitable planets Alexander James Mustill Melvyn B. Davies Anders Johansen Dynamical instability bad for habitability • Excitation of eccentricity can shift HZ or cause extreme seasons (Spiegel+10, Dressing+10) • Planets may be scattered out of HZ • Planet-planet collisions may remove biospheres, atmospheres, water • Earth-like planets may be eaten by Neptunes/Jupiters Strong dynamical effects: scattering and Kozai • Scattering: closely-spaced giant planets excite each others’ eccentricities (Chatterjee+08) • Kozai: inclined external perturber (e.g. binary) can cause very large eccentricity fluctuations (Kozai 62, Lidov 62, Naoz 16) Relevance of inner systems to HZ • If you can • form a hot Jupiter through high-eccentricity migration • damage a Kepler system at few tenths of an au • you will damage the habitable zone too Relevance of inner systems intrinsically • Large number of single-candidate systems found by Kepler relative to multiples • Is this left over from formation? Or do the multiples evolve into singles through dynamics? (Johansen+12) • Informs models of planet formation • all the Kepler systems are interestingly different to the Solar system, but do we have two interestingly different channels of planet formation or only one? What do we know about the prevalence of strong dynamical effects? • So far know little about planets in HZ • What we do know: • Violent dynamical history strong contender for hot Jupiter migration • Many giants have high -

Dynamics of the Terrestrial Planets from a Large Number of N-Body Simulations ∗ Rebecca A
Earth and Planetary Science Letters 392 (2014) 28–38 Contents lists available at ScienceDirect Earth and Planetary Science Letters www.elsevier.com/locate/epsl Dynamics of the terrestrial planets from a large number of N-body simulations ∗ Rebecca A. Fischer , Fred J. Ciesla Department of the Geophysical Sciences, University of Chicago, 5734 S Ellis Ave, Chicago, IL 60637, USA article info abstract Article history: The agglomeration of planetary embryos and planetesimals was the final stage of terrestrial planet Received 6 September 2013 formation. This process is modeled using N-body accretion simulations, whose outcomes are tested by Received in revised form 26 January 2014 comparing to observed physical and chemical Solar System properties. The outcomes of these simulations Accepted 3 February 2014 are stochastic, leading to a wide range of results, which makes it difficult at times to identify the full Availableonline25February2014 range of possible outcomes for a given dynamic environment. We ran fifty high-resolution simulations Editor: T. Elliott each with Jupiter and Saturn on circular or eccentric orbits, whereas most previous studies ran an Keywords: order of magnitude fewer. This allows us to better quantify the probabilities of matching various accretion observables, including low probability events such as Mars formation, and to search for correlations N-body simulations between properties. We produce many good Earth analogues, which provide information about the mass terrestrial planets evolution and provenance of the building blocks of the Earth. Most observables are weakly correlated Mars or uncorrelated, implying that individual evolutionary stages may reflect how the system evolved even late veneer if models do not reproduce all of the Solar System’s properties at the end. -

The History of Planet Earth
SECOND EDITION Earth’s Evolving The History of Systems Planet Earth Ronald Martin, Ph.D. University of Delaware Newark, Delaware © Jones & Bartlett Learning, LLC, an Ascend Learning Company. NOT FOR SALE OR DISTRIBUTION 9781284457162_FMxx_00i_xxii.indd 1 07/11/16 1:46 pm World Headquarters Jones & Bartlett Learning 5 Wall Street Burlington, MA 01803 978-443-5000 [email protected] www.jblearning.com Jones & Bartlett Learning books and products are available through most bookstores and online booksellers. To contact Jones & Bartlett Learning directly, call 800-832-0034, fax 978-443-8000, or visit our website, www.jblearning.com. Substantial discounts on bulk quantities of Jones & Bartlett Learning publications are available to corporations, professional associations, and other qualified organizations. For details and specific discount information, contact the special sales department at Jones & Bartlett Learning via the above contact information or send an email to [email protected]. Copyright © 2018 by Jones & Bartlett Learning, LLC, an Ascend Learning Company All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without written permission from the copyright owner. The content, statements, views, and opinions herein are the sole expression of the respective authors and not that of Jones & Bartlett Learning, LLC. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement or recommendation by Jones & Bartlett Learning, LLC, and such reference shall not be used for advertis- ing or product endorsement purposes. -

Foundations of Complex Life: Evolution, Preservation and Detection on Earth and Beyond
National Aeronautics and Space Administration NASA Astrobiology Institute Annual Science Report 2016 Team Report: Foundations of Complex Life: Evolution, Preservation and Detection on Earth and Beyond, Massachusetts Institute of Technology Foundations of Complex Life Evolution, Preservation and Detection on Earth and Beyond Lead Institution: Massachusetts Institute of Technology Team Overview Foundations of Complex Life is a research project investigating the early evolution and preservation of complex life on Earth. We seek insight into the early evolution and preservation of complex life by refining our ability to identify evidence of environmental and biological change in the late Mesoproterozoic to Neoproterozoic eras. Better understanding how signatures of life and environment are preserved will guide how and where to look for evidence for life elsewhere in the universe—directly supporting the Curiosity mission on Mars and helping set strategic goals for future explorations of the Solar System and studies of the early Earth. Our Team pursues these questions under five themes: I. The earliest history of animals: We use methods from molecular biology, experimental taphonomy, and paleontology to explore what caused the early divergence of animals. Principal Investigator: II. Paleontology, sedimentology, and geochemistry: We track the origin of complex Roger Summons protists and animals from their biologically simple origins by documenting the stratigraphy, isotopic records, and microfossil assemblages of well-preserved rock successions from 1200 to 650 million years ago. III. A preservation-induced oxygen tipping point: We investigate how changes in the preservation of organic carbon may have driven the Neoproterozoic oxygenation of the oceans coincident with the appearance of complex life. -

Nature Based Solutions FEMA
PROMOTING NATURE-BASED HAZARD MITIGATION THROUGH FEMA MITIGATION GRANTS ABBREVIATIONS ADCIRC – Advanced Circulation Model HGM Approach – Hydrogeomorphic Approach BCA – Benefit-Cost Analysis HMA – Hazard Mitigation Assistance BCR – Benefit-Cost Ratio HMGP – Hazard Mitigation Grant Program BRIC – Building Resilient Infrastructure and MSCP – Multiple Species Conservation Program Communities NBS – Nature-Based Solution C&CB – Capability- and Capacity-Building NFIP – National Flood Insurance Program CDBG-DR – Community Development Block Grant- Disaster Recovery NFWF – National Fish and Wildlife Foundation CDBG-MIT – Community Development Block NOAA – National Oceanic and Atmospheric Grant-Mitigation Administration D.C. – District of Columbia NOFO – Notice of Funding Opportunity DEM – Department of Emergency Management NPV – Net Present Value DOI – Department of the Interior SCC – State Coastal Conservancy EDYS – Ecological Dynamics Simulation SDG&E – San Diego Gas & Electric EMA – Emergency Management Agency SFHA – Special Flood Hazard Area EPA SWMM – Environmental Protection Agency SHMO – State Hazard Mitigation Officer Storm Water Management Model SLAMM – Sea Level Affecting Marshes Model FEMA – Federal Emergency Management Agency SRH-2D – Sedimentation and River Hydraulics – FIRM – Flood Insurance Rate Map Two-Dimension FMA – Flood Mitigation Assistance STWAVE – Steady-State Spectral Wave Model FMAG – Fire Management Assistance Grant TNC – The Nature Conservancy HAZUS – Hazards US USACE – U.S. Army Corps of Engineers HEC-HMS – Hydrologic -

Cosmochemistry Cosmic Background Radia�On
6/10/13 Cosmochemistry Cosmic background radiaon Dust Anja C. Andersen Niels Bohr Instute University of Copenhagen hp://www.dark-cosmology.dk/~anja Hauser & Dwek 2001 Molecule formation on dust grains Multiwavelenght MW 1 6/10/13 Gas-phase element depleons in the Concept of dust depleon interstellar medium The depleon of an element X in the ISM is defined in terms of (a logarithm of) its reducon factor below the expected abundance relave to that of hydrogen if all of the atoms were in the gas phase, [Xgas/H] = log{N(X)/N(H)} − log(X/H) which is based on the assumpon that solar abundances (X/H)are good reference values that truly reflect the underlying total abundances. In this formula, N(X) is the column density of element X and N(H) represents the column density of hydrogen in both atomic and molecular form, i.e., N(HI) + 2N(H2). The missing atoms of element X are presumed to be locked up in solids within dust grains or large molecules that are difficult to idenfy spectroscopically, with fraconal amounts (again relave to H) given by [Xgas/H] (Xdust/H) = (X/H)(1 − 10 ). Jenkins 2009 Jenkins 2009 2 6/10/13 Jenkins 2009 Jenkins 2009 The Galacc Exncon Curve Extinction curves measure the difference in emitted and observed light. Traditionally measured by comparing two stars of the same spectral type. Galactic Extinction - empirically determined: -1 -1 <A(λ)/A(V)> = a(λ ) + b(λ )/RV (Cardelli et al. 1999) • Bump at 2175 Å (4.6 µm-1) • RV : Ratio of total to selective extinction in the V band • Mean value is RV = 3.1 (blue) • Low value: RV = 1.8 (green) (Udalski 2003) • High value: RV = 5.6-5.8 (red) (Cardelli et al. -

The Solar System
5 The Solar System R. Lynne Jones, Steven R. Chesley, Paul A. Abell, Michael E. Brown, Josef Durech,ˇ Yanga R. Fern´andez,Alan W. Harris, Matt J. Holman, Zeljkoˇ Ivezi´c,R. Jedicke, Mikko Kaasalainen, Nathan A. Kaib, Zoran Kneˇzevi´c,Andrea Milani, Alex Parker, Stephen T. Ridgway, David E. Trilling, Bojan Vrˇsnak LSST will provide huge advances in our knowledge of millions of astronomical objects “close to home’”– the small bodies in our Solar System. Previous studies of these small bodies have led to dramatic changes in our understanding of the process of planet formation and evolution, and the relationship between our Solar System and other systems. Beyond providing asteroid targets for space missions or igniting popular interest in observing a new comet or learning about a new distant icy dwarf planet, these small bodies also serve as large populations of “test particles,” recording the dynamical history of the giant planets, revealing the nature of the Solar System impactor population over time, and illustrating the size distributions of planetesimals, which were the building blocks of planets. In this chapter, a brief introduction to the different populations of small bodies in the Solar System (§ 5.1) is followed by a summary of the number of objects of each population that LSST is expected to find (§ 5.2). Some of the Solar System science that LSST will address is presented through the rest of the chapter, starting with the insights into planetary formation and evolution gained through the small body population orbital distributions (§ 5.3). The effects of collisional evolution in the Main Belt and Kuiper Belt are discussed in the next two sections, along with the implications for the determination of the size distribution in the Main Belt (§ 5.4) and possibilities for identifying wide binaries and understanding the environment in the early outer Solar System in § 5.5. -

Curriculum Vitae
CURRICULUM VITAE Smadar Naoz September 2021 Contact University of California Los Angeles, Information Department of Physics & Astronomy 30 Portola Plaza, Box 951547 E-mail: [email protected] Los Angeles, CA 90095 WWW: http://www.astro.ucla.edu/∼snaoz/ Research Dynamics of planetary, stellar and black hole systems, which include formation of Hot Jupiters, Interests globular clusters, spiral structure, compact objects etc. Cosmology, structure formation in the early Universe, reionization and 21cm fluctuations. Education Tel Aviv University, Tel Aviv, Israel Ph.D. in Physics, January 2010 Hebrew University of Jerusalem, Jerusalem, Israel M.S. in Physics, Magna Cum Laude, 2004 B.S. in Physics 2002 Positions University of California, Los Angeles Associate professor since July 2019 Howard & Astrid Preston Term Chair in Astrophysics since July 2018 Assistant professor 2014-2019 Harvard Smithsonian CfA, Institute for Theory and Computation Einstein Fellow, September 2012 { June 2014 ITC Fellow, September 2011 { August 2012 Northwestern University, CIERA Gruber Fellow, September 2010 { August 2011 Postdoctoral associate in theoretical astrophysics, January 2010 { August 2010 Scholarships Helen B. Warner Prize, awarded by the American Astronomical Society, 2020 Honors and Scialog fellow, and accepted proposal, Signatures of Life in the Universe, 2020/2021 (conference Awards postponed to 2021 due to COVID-19) Career Commitment to Diversity, Equity and Inclusion Award, given by UCLA Academic Senate 2019. For other diversity awards, see xDEI. Hellman Fellows Award, awarded by Hellman Fellows Program, aimed to support the research of promising Assistant Professors who show capacity for great distinction in their research, June 2017 Multiple departmental teaching awards 2015-2019, see xTeaching, for details Sloan Research Fellowships awarded by the Alfred P. -

Cosmochemical Evidence for Astrophysical Processes During the Formation of Our Solar System
Cosmochemical evidence for astrophysical processes during the formation of our solar system Glenn J. MacPhersona,1 and Alan Bossb aDepartment of Mineral Sciences, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560; and bDepartment of Terrestrial Magnetism, Carnegie Institution of Washington, Washington, DC 20015 Edited by Mark H. Thiemens, University of California, San Diego, La Jolla, CA, and approved October 25, 2011 (received for review June 22, 2011) Through the laboratory study of ancient solar system materials such as meteorites and comet dust, we can recognize evidence for the same star-formation processes in our own solar system as those that we can observe now through telescopes in nearby star-forming regions. High temperature grains formed in the innermost region of the solar system ended up much farther out in the solar system, not only the asteroid belt but even in the comet accretion region, suggesting a huge and efficient process of mass transport. Bi-polar outflows, turbulent diffusion, and marginal gravitational instability are the likely mechanisms for this transport. The presence of short-lived radionuclides in the early solar system, especially 60Fe, 26Al, and 41Ca, requires a nearby supernova shortly before our solar system was formed, suggesting that the Sun was formed in a massive star-forming region similar to Orion or Carina. Solar system formation may have been “triggered” by ionizing radiation originating from massive O and B stars at the center of an expanding HII bubble, one of which may have later provided the supernova source for the short-lived radionuclides. Alternatively, a supernova shock wave may have simultaneously triggered the collapse and injected the short-lived radionuclides. -

Dynamical Models of Terrestrial Planet Formation
Dynamical Models of Terrestrial Planet Formation Jonathan I. Lunine, LPL, The University of Arizona, Tucson AZ USA, 85721. [email protected] David P. O’Brien, Planetary Science Institute, Tucson AZ USA 85719 Sean N. Raymond, CASA, University of Colorado, Boulder CO USA 80302 Alessandro Morbidelli, Obs. de la Coteˆ d’Azur, Nice, F-06304 France Thomas Quinn, Department of Astronomy, University of Washington, Seattle USA 98195 Amara L. Graps, SWRI, Boulder CO USA 80302 Revision submitted to Advanced Science Letters May 23, 2009 Abstract We review the problem of the formation of terrestrial planets, with particular emphasis on the interaction of dynamical and geochemical models. The lifetime of gas around stars in the process of formation is limited to a few million years based on astronomical observations, while isotopic dating of meteorites and the Earth-Moon system suggest that perhaps 50-100 million years were required for the assembly of the Earth. Therefore, much of the growth of the terrestrial planets in our own system is presumed to have taken place under largely gas-free conditions, and the physics of terrestrial planet formation is dominated by gravitational interactions and collisions. The ear- liest phase of terrestrial-planet formation involve the growth of km-sized or larger planetesimals from dust grains, followed by the accumulations of these planetesimals into ∼100 lunar- to Mars- mass bodies that are initially gravitationally isolated from one-another in a swarm of smaller plan- etesimals, but eventually grow to the point of significantly perturbing one-another. The mutual perturbations between the embryos, combined with gravitational stirring by Jupiter, lead to orbital crossings and collisions that drive the growth to Earth-sized planets on a timescale of 107 − 108 years. -
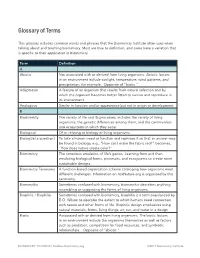
Glossary of Terms
Glossary of Terms This glossary includes common words and phrases that the Biomimicry Institute often uses when talking about and teaching biomimicry. Most are true to definition, and some have a variation that is specific to their application in biomimicry. Term Definition A Abiotic Not associated with or derived from living organisms. Abiotic factors in an environment include sunlight, temperature, wind patterns, and precipitation, for example. Opposite of "biotic." Adaptation A feature of an organism that results from natural selection and by which the organism becomes better fitted to survive and reproduce in its environment. Analogous Similar in function and/or appearance but not in origin or development. B Biodiversity The variety of life and its processes; includes the variety of living organisms, the genetic differences among them, and the communities and ecosystems in which they occur. Biological Of or relating to biology or living organisms. Biologize [a question] To take a human need or function and rephrase it so that an answer may be found in biology, e.g., "How can I make the fabric red?" becomes, "How does nature create color?" Biomimicry The conscious emulation of life’s genius. Learning from and then emulating biological forms, processes, and ecosystems to create more sustainable designs. Biomimicry Taxonomy A function-based organization scheme cataloging how organisms meet different challenges. Information on AskNature.org is organized by this taxonomy. Biomorphic Sometimes confused with biomimicry, biomorphic describes anything resembling or suggesting the forms of living organisms. Biophilic / Biophilia Sometimes confused with biomimicry, biophilia is a term popularized by E.O. Wilson to describe the extent to which humans need connection with nature and other forms of life. -

Origins of Life in the Universe Zackary Johnson
11/4/2007 Origins of Life in the Universe Zackary Johnson OCN201 Fall 2007 [email protected] Zackary Johnson http://www.soest.hawaii.edu/oceanography/zij/education.html Uniiiversity of Hawaii Department of Oceanography Class Schedule Nov‐2Originsof Life and the Universe Nov‐5 Classification of Life Nov‐7 Primary Production Nov‐9Consumers Nov‐14 Evolution: Processes (Steward) Nov‐16 Evolution: Adaptation() (Steward) Nov‐19 Marine Microbiology Nov‐21 Benthic Communities Nov‐26 Whale Falls (Smith) Nov‐28 The Marine Food Web Nov‐30 Community Ecology Dec‐3 Fisheries Dec‐5Global Ecology Dec‐12 Final Major Concepts TIMETABLE Big Bang! • Life started early, but not at the beginning, of Earth’s Milky Way (and other galaxies formed) history • Abiogenesis is the leading hypothesis to explain the beginning of life on Earth • There are many competing theories as to how this happened • Some of the details have been worked out, but most Formation of Earth have not • Abiogenesis almost certainly occurred in the ocean 20‐15 15‐94.5Today Billions of Years Before Present 1 11/4/2007 Building Blocks TIMETABLE Big Bang! • Universe is mostly hydrogen (H) and helium (He); for Milky Way (and other galaxies formed) example –the sun is 70% H, 28% He and 2% all else! Abundance) e • Most elements of interest to biology (C, N, P, O, etc.) were (Relativ 10 produced via nuclear fusion Formation of Earth Log at very high temperature reactions in large stars after Big Bang 20‐13 13‐94.7Today Atomic Number Billions of Years Before Present ORIGIN OF LIFE ON EARTH Abiogenesis: 3 stages Divine Creation 1.