UC Irvine UC Irvine Previously Published Works
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Project Report Innovation Norway
1 Project Report Innovation Norway Authors: Claudia Erber Nils-Roar Hareide 2 Distribution: Open Runde Miljøsenter AS Funded by: Rundavegen237 Innovation 6096 Runde Norway and Org. Nr. 987 410 752 MVA Runde Telephone: 70 08 08 00 Environmental Email: [email protected] Centre Web: www.rundecentre.no Date: 27.02.19 Title: Runde Miljøsenter THE DEEPSEA ENVIRONMENTAL MONITORING CLUSTER Report number: 2019-01 Authors: Number of pages: Claudia Erber (Runde Environmental Centre) 21 Nils-Roar Hareide (Runde Environmental Centre) Key words: Network, Subsea, offshore, deep-sea, biology, cluster, environment 3 1. SUMMARY This report summarizes the DeepSea Environmental Monitoring Cluster project conducted in the period from July 2018 to February 2019. The aim of the project was to consolidate the bilateral network from Brazil and Norway by participating during events in Norway and Brazil. During the events REC team tried to investigate the opportunities for new business and science during oil and gas operations. In 2018, with the GCE Subsea support, Runde Environmental Centre (REC) was granted funding from Innovation Norway for participating in many important events for the energy sector in Norway and Brazil (ONS-Offshore Northern Seas and ROG-Rio Oil and Gas). REC was invited by Innovation Norway and Finep (Brazilian Innovation Agency) to organize a multidisciplinary round-table Conference during the Norway-Brazil Week 2018, in November in Brazil. In 2019, REC participated on the GoGlobal Brazil Oil&Gas Seminar, organized by Innovation Norway at ÅKP GCE Blue Maritime, in Ålesund, Norway. During the present project, strong ties were made with Norwegian companies, as we share many common goals: access the deep-sea environment and the Brazilian oil and gas market. -

Runde Miljøsenter: North Atlantic Seabird Seminar
North Atlantic Seabird Seminar – Fosnavåg, Norway 20-21 April 2015 Michael Hundeide (ed.) Runde Environmental Centre In cooperation with: Birdlife Norway (NOF) and Norwegian Biologist Association (BiO). Distribution: Open/Closed Runde Miljøsenter AS Client(s) 6096 Runde Org. Nr. 987 410 752 MVA Telephone: +47 70 08 08 00 E-mail: [email protected] Date: Web: www.rundesenteret.no 18.09.2015 Report Runde Miljøsenter Norsk: Nordatlantisk sjøfuglseminar 2015. Fosnavåg 20. og 21. April Rapportnummer: English: North Atlantic Seabird Seminar – Fosnavåg, Norway 20-21 April 2015 Author(s): Number of pages: Michael Hundeide (ed.) 52 Key words: Seabird ecology, Runde, North Atlantic, sand eel Sammendrag (Norsk):Rapporten er en gjengivelse av de temaer som ble presentert og diskutert på det internasjonale sjøfuglseminaret i Fosnavåg 20. og 21. april 2015: En rekke sjøfuglbestander har hatt en drastisk tilbakegang på Runde i senere år. Særlig gjelder dette artene: Krykkje, havhest, toppskarv, lomvi, alke, lunde m.fl. Med unntak av havsule, storjo og havørn som har hatt en økende tendens, har så å si samtlige av de andre sjøfuglartene gått ned. Denne situasjonen er ikke enestående for Runde og Norge, samme tendens gjør seg også gjeldende i Storbritannia, på Island og i hele Nordøst -Atlanteren. Selv om årsakene til krisen hos sjøfuglene er sammensatte og komplekse, var de fleste innleggsholderne enige om følgende punkter: 1) En vesentlig del av årsaken til krisen for sjøfuglbestandene på Runde og i Nordøst-Atlanteren skyldes mangel på egnet energirik mat (fettrik fisk av rett størrelse særlig tobis i Skottland, Færøyene og Sør-Vestlige Island) i hekkesesongen. Dette kan i sin tur forklares som en indirekte effekt av en liten økning i gjennomsnittstemperatur i havet. -

Information Sheet on Ramsar Wetlands (RIS) – 2009-2012 Version
Information Sheet on Ramsar Wetlands (RIS) – 2009-2012 version Available for download from http://www.ramsar.org/ris/key_ris_index.htm. Categories approved by Recommendation 4.7 (1990), as amended by Resolution VIII.13 of the 8th Conference of the Contracting Parties (2002) and Resolutions IX.1 Annex B, IX.6, IX.21 and IX. 22 of the 9th Conference of the Contracting Parties (2005). Notes for compilers: 1. The RIS should be completed in accordance with the attached Explanatory Notes and Guidelines for completing the Information Sheet on Ramsar Wetlands. Compilers are strongly advised to read this guidance before filling in the RIS. 2. Further information and guidance in support of Ramsar site designations are provided in the Strategic Framework and guidelines for the future development of the List of Wetlands of International Importance (Ramsar Wise Use Handbook 14, 3rd edition). A 4th edition of the Handbook is in preparation and will be available in 2009. 3. Once completed, the RIS (and accompanying map(s)) should be submitted to the Ramsar Secretariat. Compilers should provide an electronic (MS Word) copy of the RIS and, where possible, digital copies of all maps. 1. Name and address of the compiler of this form: FOR OFFICE USE ONLY. DD MM YY Maria Aastum, County Governor of Møre og Romsdal, Fylkeshuset, 6404 Molde Designation date Site Reference Number 2. Date this sheet was completed/updated: May 2013 3. Country: Norway 4. Name of the Ramsar site: The precise name of the designated site in one of the three official languages (English, French or Spanish) of the Convention. -

Important Bird Areas and Potential Ramsar Sites in Europe
cover def. 25-09-2001 14:23 Pagina 1 BirdLife in Europe In Europe, the BirdLife International Partnership works in more than 40 countries. Important Bird Areas ALBANIA and potential Ramsar Sites ANDORRA AUSTRIA BELARUS in Europe BELGIUM BULGARIA CROATIA CZECH REPUBLIC DENMARK ESTONIA FAROE ISLANDS FINLAND FRANCE GERMANY GIBRALTAR GREECE HUNGARY ICELAND IRELAND ISRAEL ITALY LATVIA LIECHTENSTEIN LITHUANIA LUXEMBOURG MACEDONIA MALTA NETHERLANDS NORWAY POLAND PORTUGAL ROMANIA RUSSIA SLOVAKIA SLOVENIA SPAIN SWEDEN SWITZERLAND TURKEY UKRAINE UK The European IBA Programme is coordinated by the European Division of BirdLife International. For further information please contact: BirdLife International, Droevendaalsesteeg 3a, PO Box 127, 6700 AC Wageningen, The Netherlands Telephone: +31 317 47 88 31, Fax: +31 317 47 88 44, Email: [email protected], Internet: www.birdlife.org.uk This report has been produced with the support of: Printed on environmentally friendly paper What is BirdLife International? BirdLife International is a Partnership of non-governmental conservation organisations with a special focus on birds. The BirdLife Partnership works together on shared priorities, policies and programmes of conservation action, exchanging skills, achievements and information, and so growing in ability, authority and influence. Each Partner represents a unique geographic area or territory (most often a country). In addition to Partners, BirdLife has Representatives and a flexible system of Working Groups (including some bird Specialist Groups shared with Wetlands International and/or the Species Survival Commission (SSC) of the World Conservation Union (IUCN)), each with specific roles and responsibilities. I What is the purpose of BirdLife International? – Mission Statement The BirdLife International Partnership strives to conserve birds, their habitats and global biodiversity, working with people towards sustainability in the use of natural resources. -
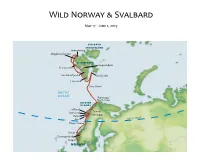
2019 Wild Norway & Svalbard Field Report
Wild Norway & Svalbard May 17 - June 2, 2019 SVALBARD ARCHIPELAGO Smeerenburg Magdalenefjorden SPITSBERGEN Longyearbyen Poolepynten Van Mijenfjorden Storfjorden Hornsund Bear Island ARCTIC OCEAN Skarsvaag/ North Cape LOFOTEN ISLANDS Trollfjord Stamsund Tromsø Kjerringøy Reine CLE Røst ARCTIC CIR Husey/Sanna Runde Geirangerfjord Bergen NORWAY Sunday, May 19, 2019 Bergen, Norway / Embark Ocean Adventurer Unusual for Bergen, it was a dry, warm, and sunny weekend when we arrived here to begin our travels in Wild Norway and Svalbard. Bergen experiences only five rain-free days a year, and this exceptional weather was being fully enjoyed by the locals, especially as this was also the Norwegian National Day holiday weekend. We set out on foot early this morning, for a tour of the old wharf-side district of Bryggen with its picturesque and higgledy-piggledy medieval wooden houses, their colorful gables lit up by today’s bright sunshine. Next, we boarded coaches which took us to Troldhaugen, the home of Norwegian composer Edvard Grieg for over 20 years. The hut where he did much of his work still stands on the lakeside at the foot of the garden. We were treated to an excellent piano recital of some of Grieg’s music before we went to lunch. All too soon, it was time to return to Bergen where our home for the next two weeks, the Ocean Adventurer, awaited us. Once settled in our cabins, our Expedition Leader, John Yersin, introduced us to his team of Zodiac drivers and lecturers who would be accompanying us on our journey over the next two weeks. -

Norway Ramsar Information Sheet Published on 17 April 2018 Update Version, Previously Published on : 27 May 2013
RIS for Site no. 2164, Runde, Norway Ramsar Information Sheet Published on 17 April 2018 Update version, previously published on : 27 May 2013 Norway Runde Designation date 27 May 2013 Site number 2164 Coordinates 62°24'33"N 05°39'54"E Area 351,00 ha https://rsis.ramsar.org/ris/2164 Created by RSIS V.1.6 on - 18 May 2020 RIS for Site no. 2164, Runde, Norway Color codes Fields back-shaded in light blue relate to data and information required only for RIS updates. Note that some fields concerning aspects of Part 3, the Ecological Character Description of the RIS (tinted in purple), are not expected to be completed as part of a standard RIS, but are included for completeness so as to provide the requested consistency between the RIS and the format of a ‘full’ Ecological Character Description, as adopted in Resolution X.15 (2008). If a Contracting Party does have information available that is relevant to these fields (for example from a national format Ecological Character Description) it may, if it wishes to, include information in these additional fields. 1 - Summary Summary The Site is located on the northwest coast of Norway and it consists of several protected areas on the island Runde, as well as the neighbouring island Grasøyane . Island Runde is exposed to the Norwegian Sea and includes large parts of the western coastline protected as bird reserves, and an inland mire area protected as a nature reserve, Goksøyrmyrane . The mountains rise steeply around the island Runde, and the island is the southernmost and third largest bird cliff nesting site in Norway. -

The Status and Trends of Seabirds Breeding in Norway and Svalbard
The status and trends of seabirds breeding in Norway and Svalbard Per Fauchald, Tycho Anker-Nilssen, Robert T. Barrett, Jan Ove Bust- nes, Bård-Jørgen Bårdsen, Signe Christensen-Dalsgaard, Sébastien Descamps, Sigrid Engen, Kjell Einar Erikstad, Sveinn Are Hanssen, Svein-Håkon Lorentsen, Børge Moe, Tone K. Reiertsen, Hallvard Strøm, Geir Helge Systad NINA Publications NINA Report (NINA Rapport) This is a electronic series beginning in 2005, which replaces the earlier series NINA commissioned reports and NINA project reports. This will be NINA’s usual form of reporting completed research, monitoring or review work to clients. In addition, the series will include much of the institute’s other reporting, for example from seminars and conferences, results of internal research and review work and literature studies, etc. NINA report may also be issued in a second language where appropri- ate. NINA Special Report (NINA Temahefte) As the name suggests, special reports deal with special subjects. Special reports are produced as required and the series ranges widely: from systematic identification keys to information on im- portant problem areas in society. NINA special reports are usually given a popular scientific form with more weight on illustrations than a NINA report. NINA Factsheet (NINA Fakta) Factsheets have as their goal to make NINA’s research results quickly and easily accessible to the general public. The are sent to the press, civil society organisations, nature management at all lev- els, politicians, and other special interests. Fact sheets give a short presentation of some of our most important research themes. Other publishing In addition to reporting in NINA’s own series, the institute’s employees publish a large proportion of their scientific results in international journals, popular science books and magazines. -
Wild Norway & Svalbard
WILD NORWAY & SVALBARD May 17 – June 2, 2019 | 17 Days | Aboard the Ocean Adventurer SVALBARD ARCHIPELAGO Moffen Island Wildlife Highlights NORDVEST-SPITSBERGEN NATIONAL PARK • Explore the Norwegian mainland for Monaco Glacier birds of forest and field including SPITSBERGEN European Golden-Plover, Redwing, Longyearbyen Poolepynten Ring Ouzel, and more. Farmhamna • Discover hundreds of thousands of SOR-SPITSBERGEN NATIONAL PARK breeding seabirds including Black- legged Kittiwakes, Black Guillemots, Bear Island Thick-billed Murres, Dovekies, and Ivory Gulls. ARCTIC OCEAN Skarsvaag/ North Cape • Search for the Svalbard Archipelago’s LOFOTEN legendary wildlife: Walrus, seals, Arctic ISLANDS Fox, and the ultimate prize—Polar Trollfjord Bears! Stamsund Tromsø Kjerringøy Reine CLE Røst ARCTIC CIR • Watch for Minke and Fin whales and Expedition Stop beautiful Orcas. Runde Geiranger • Itinerary .................................... page 2 > • Flight Information ...................... page 3 > Bergen Oslo • Ship, Deck Plan & Rates ............ page 4 > NORWAY • Featured Leader ........................ page 5 > • Know Before You Go ................. page 5 > POOLEPYNTEN BEAR ISLAND DOVEKIE ©Rich Price ©Rick Price Shags, and Guillemots. From Zodiacs look for the seals that rest Itinerary on some of the smaller offshore islands. This afternoon we sail the western coast of Norway, weaving among the dramatic fjords with their verdant slopes and towering cliffs. Based on the expeditionary nature of our trips, there may be ongoing enhancements to this itinerary. Wednesday, May 22 EXPEDITION STOP Friday, May 17, 2019 Weather permitting, board Zodiacs and explore the rugged DEPART USA coastline for shorebirds, Willow Warbler, wagtails, and pipits. Board your independent flight to Norway. Thursday, May 23 Saturday, May 18 KJERRINGØY / RØST, LOFOTEN ISLANDS Officially north of the Arctic Circle, the 19th-century trading station BERGEN, NORWAY Arrive in Bergen this afternoon and transfer to our hotel. -

Cradle to Cradle Islands Runde And
cradle to cradle islands “Dear reader The Cradle to Cradle Islands project can look back on a In September we joined the 400 year celebration of successful first project year. Ideas together with the the friendship between the United States and The research have been translated into projects. In October Netherlands, which gives us lots of opportunities. The Cradle to Cradle we had our second transnational partner meeting on On behalf of the team of Cradle to Cradle Islands Islands project Runde Island in Norway. What a creativity and enthu- We wish you a happy 2010.” siasm! Results of the investigation phase have been works towards a presented and shared with partners. Workshops on Anne de Vries, project manager clean and healthy the themes of Water, Energy and Materials have been Hans van Meerendonk, project coordinator future for the islands conducted. Ideas for potential pilot projects have been in the North Sea discussed and agreed that before the end of the year region through the pilots from each partner will be selected. We had implementation of two productive days on Runde where we got to know innovations based each other better. The clusters Water/Energy, Energy/ on the Cradle to Mobility and Materials/Energy are well on track to get a Cradle® concept. good perception. You can find more information about the progress of each cluster further in the newsletter. Construction of Runde Environmental Centre and the Cradle to Cradle® concept Runde and Cradle to Cradle While the centre’s main focus is to provide facilities • To limit heat loss across exterior surfaces, walls in the for environmental research and education on the three-section construction (apartment complex, visitor Norwegian west coast Island of Runde, the testing centre and office/laboratory wing) have 30cm rock and demonstration of innovative and ’green’ wool insulation, while the roof has 45cm. -

Law and Practice Regarding Coin Finds
LAW AND PRACTICE REGARDING COIN FINDS Svein H. Gullbekk & Håkon Roland THE LAW AND PRACTICE REGARDING COIN FINDS: NORWAY The Heritage Conservation Act in Norway was introduced as late as 1905. The law had been in preparation for years, but was first statutory by Parliament after Norway became independent from Sweden earlier that year. The history of the heritage administration in Norway was to a large degree determined by political unions with Denmark from 1397 to 1814 and Sweden from 1814 to 1905. Even though a Norwegian Constitution was introduced in 1814 and the interest in heritage issues was prolific in the beginning of the nineteenth century, laws and regulation of antiqua- rian issues did not become statutory before the early 20th century. Cultural heritage in the form of precious metal and coins buried in the ground has long been a feature of Norwegian society. The earliest references to such treasures are found within saga literature. The most famous example is from Egilsoga (Egil’s Saga), in which the archetypal Viking Egil is shown as an old man burying the treasure of coins that he had received from King Athelstan many years earlier after the battle of Brunanburh in 937.[1] We know also that hoards of Roman coins were discovered in the Viking or early medieval period as these Roman coins were then reused as currency, and we have found them with Viking and medieval currency and other objects in different contexts. The earliest Norwegian laws on buried treasure can be seen in the Gula- thing of Western Norway’s Landscape Law. -

Urban Gardeners in Oslo: an Explorative Study of Their Motivations and the Perceived Effects of Gardening on Quality of Life
Master’s Thesis 2019 30 ECTS BIOVIT Urban gardeners in Oslo: An explorative study of their motivations and the perceived effects of gardening on quality of life Carolina Hernandez Lasso & Rina Dula Shabi MSc Agroecology ACKNOWLEDGEMENTS We would like to thank the Agroecology program and the Faculty of Biosciences for allowing us to do an interdisciplinary research between master programs of International Environmental Studies and Agroecology. Thanks to all the professors who have contributed during our development as master students, as well as to Cathrine Strømø and for always being there as a student advisor. This master thesis would not have happened without the Cultivating Public Spaces research project. A big thanks to the entire team for welcoming us and allowing us to collaborate. A special thank you to Anna Marie Nicolaysen, Vebjørn Egner Stafseng and Katinka Horgen Evensen for your insight, support and advice. To all the gardening initiatives and their organizers and project leaders, thank you for giving us the opportunity to visit the gardens, allowing us to get involved and for your generosity. We are beyond grateful for all the participants involved for their time and interest in our research. We would also like to thank our families and friends for your moral and scholar contribution and always supporting us through our ups and downs. Especially to our partners, Magnus and Esli for your patience and keeping us motivated throughout the whole process. Lastly, we must extend our appreciation to Anna Marie Nicolaysen and Erik Gomez-Baggethun for your thoughtful feedback and guidance until the very last part of this research process. -

Runde Marine Station
Runde Marine Station A new marine research station at Runde, Norway Prestudy on project development, science networking and market Report (abbreviated web-version) to NORA Møre og Romsdal Nordisk Fylkeskommune HERØY KOMMUNE Atlandsamarbejd Post-boks 259, FO-10 Torshavn, Færøyane 6404 Molde, Norge 6090 Fosnavåg, Norge Prepared by the Runde Group Runde/Ulsteinvik, Norway, 1st October, 2004 Runde Marine Station -report to NORA, 1st October, 2004. © Runde Group 2004 Preface The island Runde on the west coast of Norway has for a long time attracted regular tourists as well as scientists interested in the natural environment on and around the island. On this background, discussions were initiated in the 1990’s to establish a field station on the island that could provide services to the scientists as well as to other visitors and activities. A local group of people (the Runde Group) occupied with this idea has worked informally on planning the station for several years. Then, in March 2003, an application for a pilot study was submitted to the Nordic Council NORA (North Atlantic Cooperation) programme with the aim to integrate Runde in a Nordic/West-Nordic network cooperation among other stations in this region with a similar scope. The application was accepted in June 2003, with a tentative project time frame of 1 year. NORA underscored their interest to get the West-Nordic perspective considered. The municipality of Herøy to which Runde belongs, provided matching funding that otherwise was covered by the work of the project group itself. The present report summarises the work performed during the project period, which according to the scope focussed on networking and science.