Clostridium Botulinum Strain Hall a and Comparative Analysis of the Clostridial Genomes Mohammed Sebaihia,1 Michael W
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Shelf-Stable Food Safety
United States Department of Agriculture Food Safety and Inspection Service Food Safety Information PhotoDisc Shelf-Stable Food Safety ver since man was a hunter-gatherer, he has sought ways to preserve food safely. People living in cold climates Elearned to freeze food for future use, and after electricity was invented, freezers and refrigerators kept food safe. But except for drying, packing in sugar syrup, or salting, keeping perishable food safe without refrigeration is a truly modern invention. What does “shelf stable” Foods that can be safely stored at room temperature, or “on the shelf,” mean? are called “shelf stable.” These non-perishable products include jerky, country hams, canned and bottled foods, rice, pasta, flour, sugar, spices, oils, and foods processed in aseptic or retort packages and other products that do not require refrigeration until after opening. Not all canned goods are shelf stable. Some canned food, such as some canned ham and seafood, are not safe at room temperature. These will be labeled “Keep Refrigerated.” How are foods made In order to be shelf stable, perishable food must be treated by heat and/ shelf stable? or dried to destroy foodborne microorganisms that can cause illness or spoil food. Food can be packaged in sterile, airtight containers. All foods eventually spoil if not preserved. CANNED FOODS What is the history of Napoleon is considered “the father” of canning. He offered 12,000 French canning? francs to anyone who could find a way to prevent military food supplies from spoiling. Napoleon himself presented the prize in 1795 to chef Nicholas Appert, who invented the process of packing meat and poultry in glass bottles, corking them, and submerging them in boiling water. -

Botulism Fact Sheet
Botulism Fact Sheet 1. What is botulism? - Botulism is a rare but serious paralytic illness caused by a nerve toxin (poison) produced by the bacterium Clostridium botulinum. There are three main kinds of botulism. (a) Foodborne botulism is caused by eating foods that contain the botulism toxin. (b) Wound botulism is caused by toxin produced from a wound infected with Clostridium botulinum. (c) Infant botulism is caused when Clostridium botulinum bacteria grow in the immature infant intestines and release toxin. All forms of botulism can be fatal and are considered medical emergencies. Foodborne botulism can be especially dangerous because widespread illness can occur from eating a contaminated food. 2. What is Clostridium botulinum? - Clostridium botulinum belongs to is a group of bacteria commonly found in soil. They grow best in low oxygen conditions. The bacteria form spores which allow them to survive in a dormant state in the soil until exposed to conditions that can support their growth. In low oxygen conditions, the growing bacteria release botulinum toxin. There are seven types of botulism toxin designated by the letters A through G; only types A, B, E and F cause illness in humans. 3. How common is botulism? - In the United States an average of 45 cases are reported each year. Of these, approximately 15% are foodborne, 65% are infant botulism, and the rest are wound botulism. Outbreaks of foodborne botulism involving two or more persons occur most years and usually are caused by eating improperly processed home-canned foods. The number of Foodborne and Infant botulism cases has changed little in recent years, but wound botulism has increased because of the use of black-tar heroin. -

National Enteric Disease Surveillance: Botulism
Natio nal Enteric Disease Surveillance: Botulism Surveillance Overview Surveillance System Overview: National Botulism Surveillance System Botulism is a rare but serious paralytic illness caused by a nerve toxin that is produced by the bacterium Clostridium botulinum and sometimes by strains of Clostridium butyricum and Clostridium baratii. Botulism can be treated with an antitoxin that blocks the action of toxin circulating in the blood. Antitoxin for children one year of age and older and for adults is available through the Centers for Disease Control and Prevention (CDC), the Alaska Division of Public Health (ADPH), and the California Department of Public Health (CDPH); Colored transmission electron micrograph of the antitoxin for infants is available from CDPH. Gram-positive anaerobic bacteria Clostridium botulinum Antitoxin can be released through state public health officials for suspected botulism cases and is most effective when administered early in a patient’s illness. State public health officials can reach the CDC clinical emergency botulism service for consultation and antitoxin 24/7 at 770-488-7100. Physicians should contact their state health department as soon as they suspect that a patient may have botulism. For surveillance purposes, CDC categorizes human botulism cases into four transmission categories: foodborne, wound, infant, and other. Foodborne botulism is caused by the consumption of foods containing pre-formed botulinum toxin. Wound botulism is caused by toxin produced in a wound infected with Clostridium botulinum. Infant botulism by definition occurs in persons less than one year of age and is caused by consumption of spores of C. botulinum, which then grow and release toxins in the intestines. -

Role of Hydrogen Peroxide Vapor (HPV) for the Disinfection of Hospital Surfaces Contaminated by Multiresistant Bacteria
pathogens Review Role of Hydrogen Peroxide Vapor (HPV) for the Disinfection of Hospital Surfaces Contaminated by Multiresistant Bacteria Michele Totaro, Beatrice Casini , Sara Profeti, Benedetta Tuvo, Gaetano Privitera and Angelo Baggiani * Department of Translational Research and the New Technologies in Medicine and Surgery, University of Pisa, 56123 Pisa, Italy; [email protected] (M.T.); [email protected] (B.C.); [email protected] (S.P.); [email protected] (B.T.); [email protected] (G.P.) * Correspondence: [email protected]; Tel.: 050-221-3583; Fax: 050-221-3588 Received: 10 April 2020; Accepted: 22 May 2020; Published: 24 May 2020 Abstract: The emergence of multiresistant bacterial strains as agents of healthcare-related infection in hospitals has prompted a review of the control techniques, with an added emphasis on preventive measures, namely good clinical practices, antimicrobial stewardship, and appropriate environmental cleaning. The latter item is about the choice of an appropriate disinfectant as a critical role due to the difficulties often encountered in obtaining a complete eradication of environmental contaminations and reservoirs of pathogens. The present review is focused on the effectiveness of hydrogen peroxide vapor, among the new environmental disinfectants that have been adopted. The method is based on a critical review of the available literature on this topic Keywords: hydrogen peroxide vapor; multidrug-resistant bacteria; hospital disinfection 1. Introduction The disinfection of hospital surfaces is a complex operation aimed at reducing the pathogenic microorganism load. An ideal disinfectant must be safe for human health. It may have a good stability in the environment and may be free of toxic activity [1–4]. -

Botulism Guide for Health Care Professionals
Botulism Guide for Health Care Professionals This information requires knowledgeable interpretation and is intended primarily for use by health care workers and facilities/organizations providing health care including pharmacies, hospitals, long-term care homes, community-based health care service providers and pre-hospital emergency services. Population and Public Health Division Ministry of Health and Long-Term Care March 2017 AT A GLANCE A Quick Response Guide to Botulism Botulism – The treatment of botulism is guided by clinical diagnosis The initial diagnosis of botulism should be based on a history of recent exposure, consistent clinical symptoms and elimination of other illnesses in the differential. Treatment should not wait for laboratory confirmation. All treatment and management decisions should be made based on clinical diagnosis. Initial Presentation and evaluation of signs and symptoms There are several clinically distinct forms of botulism. All forms produce the same neurological signs and symptoms of symmetrical cranial nerve palsies followed by descending, symmetric flaccid paralysis of voluntary muscles, which may progress to respiratory compromise and death. Additional symptoms (e.g., gastrointestinal signs in foodborne cases) may also be seen in some forms. Read more on the disease on page 2 Reading the section on Differential Diagnosis and the referenced articles will assist with making the diagnosis of botulism – you will find this on page 3 Place a request for Botulinum Antitoxin (BAT) or BabyBIG® Ministry of Health and Long-Term Care (ministry) staff will arrange for the shipment of BAT. Information on ordering BAT and BabyBIG (BabyBIG has a different ordering process) is on page 5 Laboratory Diagnosis and Specimen Collection Clinical specimens must be obtained prior to administering treatment with botulinum antitoxin. -

Clostridium Amazonitimonense, Clostridium Me
ORIGINAL ARTICLE Taxonogenomic description of four new Clostridium species isolated from human gut: ‘Clostridium amazonitimonense’, ‘Clostridium merdae’, ‘Clostridium massilidielmoense’ and ‘Clostridium nigeriense’ M. T. Alou1, S. Ndongo1, L. Frégère1, N. Labas1, C. Andrieu1, M. Richez1, C. Couderc1, J.-P. Baudoin1, J. Abrahão2, S. Brah3, A. Diallo1,4, C. Sokhna1,4, N. Cassir1, B. La Scola1, F. Cadoret1 and D. Raoult1,5 1) Aix-Marseille Université, Unité de Recherche sur les Maladies Infectieuses et Tropicales Emergentes, UM63, CNRS 7278, IRD 198, INSERM 1095, Marseille, France, 2) Laboratório de Vírus, Departamento de Microbiologia, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 3) Hopital National de Niamey, BP 247, Niamey, Niger, 4) Campus Commun UCAD-IRD of Hann, Route des pères Maristes, Hann Maristes, BP 1386, CP 18524, Dakar, Senegal and 5) Special Infectious Agents Unit, King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia Abstract Culturomics investigates microbial diversity of the human microbiome by combining diversified culture conditions, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and 16S rRNA gene identification. The present study allowed identification of four putative new Clostridium sensu stricto species: ‘Clostridium amazonitimonense’ strain LF2T, ‘Clostridium massilidielmoense’ strain MT26T, ‘Clostridium nigeriense’ strain Marseille-P2414T and ‘Clostridium merdae’ strain Marseille-P2953T, which we describe using the concept of taxonogenomics. We describe the main characteristics of each bacterium and present their complete genome sequence and annotation. © 2017 Published by Elsevier Ltd. Keywords: ‘Clostridium amazonitimonense’, ‘Clostridium massilidielmoense’, ‘Clostridium merdae’, ‘Clostridium nigeriense’, culturomics, emerging bacteria, human microbiota, taxonogenomics Original Submission: 18 August 2017; Revised Submission: 9 November 2017; Accepted: 16 November 2017 Article published online: 22 November 2017 intestine [1,4–6]. -

Botulism Manual
Preface This report, which updates handbooks issued in 1969, 1973, and 1979, reviews the epidemiology of botulism in the United States since 1899, the problems of clinical and laboratory diagnosis, and the current concepts of treatment. It was written in response to a need for a comprehensive and current working manual for epidemiologists, clinicians, and laboratory workers. We acknowledge the contributions in the preparation of this review of past and present physicians, veterinarians, and staff of the Foodborne and Diarrheal Diseases Branch, Division of Bacterial and Mycotic Diseases (DBMD), National Center for Infectious Diseases (NCID). The excellent review of Drs. K.F. Meyer and B. Eddie, "Fifty Years of Botulism in the United States,"1 is the source of all statistical information for 1899-1949. Data for 1950-1996 are derived from outbreaks reported to CDC. Suggested citation Centers for Disease Control and Prevention: Botulism in the United States, 1899-1996. Handbook for Epidemiologists, Clinicians, and Laboratory Workers, Atlanta, GA. Centers for Disease Control and Prevention, 1998. 1 Meyer KF, Eddie B. Fifty years of botulism in the U.S. and Canada. George Williams Hooper Foundation, University of California, San Francisco, 1950. 1 Dedication This handbook is dedicated to Dr. Charles Hatheway (1932-1998), who served as Chief of the National Botulism Surveillance and Reference Laboratory at CDC from 1975 to 1997. Dr. Hatheway devoted his professional life to the study of botulism; his depth of knowledge and scientific integrity were known worldwide. He was a true humanitarian and served as mentor and friend to countless epidemiologists, research scientists, students, and laboratory workers. -
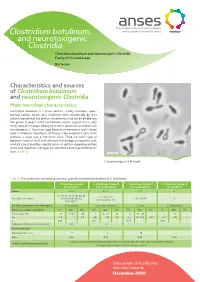
Clostridium Botulinum, and Neurotoxigenic Clostridia Clostridium Botulinum and Neurotoxigenic Clostridia Family of Clostridiaceae Bacterium
Clostridium botulinum, and neurotoxigenic Clostridia Clostridium botulinum and neurotoxigenic Clostridia Family of Clostridiaceae Bacterium Characteristics and sources of Clostridium botulinum and neurotoxigenic Clostridia Main microbial characteristics Clostridium botulinum is a Gram-positive, strictly anaerobic, spore- forming bacillus. Strains of C. botulinum differ considerably by their cultural, biochemical and genetic characteristics and can be divided into four groups (Groups I to IV). Furthermore, certain atypical strains, only rarely isolated in Europe, belonging to other species of Clostridium, are neurotoxigenic: C. butyricum (type E botulinum neurotoxin) and C. baratii (type F botulinum neurotoxin). With only a few exceptions, each strain produces a single type of botulinum toxin. There are seven types of botulinum toxin (A to G) with different immunological properties, each of which is neutralised by a specific serum. In addition, depending on their amino acid sequences, sub-types are identified in each type of botulinum toxin (Table 1). C. botulinum type A. © M. Popoff Table 1. Characteristics concerning survival, growth and toxin production of C. botulinum C. botulinum Group I C. botulinum Group II C. botulinum Group III C. botulinum Group IV (proteolytic) (non-proteolytic) (non-proteolytic) (proteolytic) Toxins A, B, F B, E, F C, D G A1, A2, A3, A4, A5, B1, B2, B3, E1, E2, E3, E6, Sous-types de toxines bivalent B (Ba, Bf, Ab), C, D, C/D, D/C G non-proteolytic B, F proteolytic F Bactéries apparentées non toxinogènes C. sporogenes Pas de nom d’espèces C. novyi C. subterminale Croissance cellules végétatives Min. Opt. Max. Min. Opt. Max. Min. Opt. Max. Min. -

Fimbriae Porphyromonas Gingivalis
TLR2 Transmodulates Monocyte Adhesion and Transmigration via Rac1- and PI3K-Mediated Inside-Out Signaling in Response to Porphyromonas gingivalis This information is current as Fimbriae of September 25, 2021. Evlambia Harokopakis, Mohamad H. Albzreh, Michael H. Martin and George Hajishengallis J Immunol 2006; 176:7645-7656; ; doi: 10.4049/jimmunol.176.12.7645 Downloaded from http://www.jimmunol.org/content/176/12/7645 References This article cites 58 articles, 32 of which you can access for free at: http://www.jimmunol.org/ http://www.jimmunol.org/content/176/12/7645.full#ref-list-1 Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists by guest on September 25, 2021 • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2006 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology TLR2 Transmodulates Monocyte Adhesion and Transmigration via Rac1- and PI3K-Mediated Inside-Out Signaling in Response to Porphyromonas gingivalis Fimbriae Evlambia Harokopakis,* Mohamad H. Albzreh,‡ Michael H. Martin,*† and George Hajishengallis2*† We present evidence for a novel TLR2 function in transmodulating the adhesive activities of human monocytes in response to the fimbriae of Porphyromonas gingivalis, a pathogen implicated in chronic periodontitis and atherosclerosis. -

Identification and Antimicrobial Susceptibility Testing of Anaerobic
antibiotics Review Identification and Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Rubik’s Cube of Clinical Microbiology? Márió Gajdács 1,*, Gabriella Spengler 1 and Edit Urbán 2 1 Department of Medical Microbiology and Immunobiology, Faculty of Medicine, University of Szeged, 6720 Szeged, Hungary; [email protected] 2 Institute of Clinical Microbiology, Faculty of Medicine, University of Szeged, 6725 Szeged, Hungary; [email protected] * Correspondence: [email protected]; Tel.: +36-62-342-843 Academic Editor: Leonard Amaral Received: 28 September 2017; Accepted: 3 November 2017; Published: 7 November 2017 Abstract: Anaerobic bacteria have pivotal roles in the microbiota of humans and they are significant infectious agents involved in many pathological processes, both in immunocompetent and immunocompromised individuals. Their isolation, cultivation and correct identification differs significantly from the workup of aerobic species, although the use of new technologies (e.g., matrix-assisted laser desorption/ionization time-of-flight mass spectrometry, whole genome sequencing) changed anaerobic diagnostics dramatically. In the past, antimicrobial susceptibility of these microorganisms showed predictable patterns and empirical therapy could be safely administered but recently a steady and clear increase in the resistance for several important drugs (β-lactams, clindamycin) has been observed worldwide. For this reason, antimicrobial susceptibility testing of anaerobic isolates for surveillance -

Clostridium Botulinum1 Keith R
FSHN0406 Preventing Foodborne Illness: Clostridium botulinum1 Keith R. Schneider, Renée M. Goodrich Schneider, Ploy Kurdmongkoltham, and Bruna Bertoldi2 This fact sheet is part of a series that discusses foodborne potent neurotoxin that causes botulism, a serious paralytic pathogens of interest to food handlers, processors, retailers, condition that can lead to death. and consumers. There are seven types of C. botulinum (A, B, C, D, E, F, and What is Clostridium botulinum? G), each distinguished by the production of serologically distinct toxins. Of the seven types, A, B, E, and rarely F Clostridium botulinum is the bacterium that causes can cause botulism in humans, while types C and D cause botulism. Clostridium botulinum is a Gram-positive, slightly botulism in animals and birds. Type G was identified in curved, motile, anaerobic, rod-shaped bacterium that 1970 but has not been determined as a cause of botulism in produces heat-resistant endospores. These endospores, humans or animals (FDA 2012; Sobel 2005). which are very resistant to a number of environmental stresses, such as heat and high acid, can become activated in anaerobic environments, low acidity (pH > 4.6), high How is Clostridium botulinum moisture content, and in temperatures ranging from 40°F transmitted? to 250°F (4°C to 121°C) (Sobel et al. 2004). In hostile The CDC categorizes human botulism cases into five environmental conditions, the heat-resistant spores enable transmission categories: foodborne, infant, wound, adult the bacteria to survive for extended periods of time in a intestinal toxemia, and iatrogenic botulism. (CDC 2017a). dormant state until conditions become more favorable. -

The Gram Positive Bacilli of Medical Importance Chapter 19
The Gram Positive Bacilli of Medical Importance Chapter 19 MCB 2010 Palm Beach State College Professor Tcherina Duncombe Medically Important Gram-Positive Bacilli 3 General Groups • Endospore-formers: Bacillus, Clostridium • Non-endospore- formers: Listeria • Irregular shaped and staining properties: Corynebacterium, Proprionibacterium, Mycobacterium, Actinomyces 3 General Characteristics Genus Bacillus • Gram-positive/endospore-forming, motile rods • Mostly saprobic • Aerobic/catalase positive • Versatile in degrading complex macromolecules • Source of antibiotics • Primary habitat:soil • 2 species of medical importance: – Bacillus anthracis right – Bacillus cereus left 4 Bacillus anthracis • Large, block-shaped rods • Central spores: develop under all conditions except in the living body • Virulence factors – polypeptide capsule/exotoxins • 3 types of anthrax: – cutaneous – spores enter through skin, black sore- eschar; least dangerous – pulmonary –inhalation of spores – gastrointestinal – ingested spores Treatment: penicillin, tetracycline Vaccines (phage 5 sensitive) 5 Bacillus cereus • Common airborne /dustborne; usual methods of disinfection/ antisepsis: ineffective • Grows in foods, spores survive cooking/ reheating • Ingestion of toxin-containing food causes nausea, vomiting, abdominal cramps, diarrhea; 24 hour duration • No treatment • Increasingly reported in immunosuppressed article 6 Genus Clostridium • Gram-positive, spore-forming rods • Obligate Anaerobes • Catalase negative • Oval or spherical spores • Synthesize organic