Ecological Factors Rather Than Temporal Factors Dominate the Evolution Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distritos Declarados Zona Catastrada.Xlsx
Distritos de Zona Catastrada "zona 1" 1-San José 2-Alajuela3-Cartago 4-Heredia 5-Guanacaste 6-Puntarenas 7-Limón 104-PURISCAL 202-SAN RAMON 301-Cartago 304-Jiménez 401-Heredia 405-San Rafael 501-Liberia 508-Tilarán 601-Puntarenas 705- Matina 10409-CHIRES 20212-ZAPOTAL 30101-ORIENTAL 30401-JUAN VIÑAS 40101-HEREDIA 40501-SAN RAFAEL 50104-NACASCOLO 50801-TILARAN 60101-PUNTARENAS 70501-MATINA 10407-DESAMPARADITOS 203-Grecia 30102-OCCIDENTAL 30402-TUCURRIQUE 40102-MERCEDES 40502-SAN JOSECITO 502-Nicoya 50802-QUEBRADA GRANDE 60102-PITAHAYA 703-Siquirres 106-Aserri 20301-GRECIA 30103-CARMEN 30403-PEJIBAYE 40104-ULLOA 40503-SANTIAGO 50202-MANSIÓN 50803-TRONADORA 60103-CHOMES 70302-PACUARITO 10606-MONTERREY 20302-SAN ISIDRO 30104-SAN NICOLÁS 306-Alvarado 402-Barva 40504-ÁNGELES 50203-SAN ANTONIO 50804-SANTA ROSA 60106-MANZANILLO 70307-REVENTAZON 118-Curridabat 20303-SAN JOSE 30105-AGUACALIENTE O SAN FRANCISCO 30601-PACAYAS 40201-BARVA 40505-CONCEPCIÓN 50204-QUEBRADA HONDA 50805-LIBANO 60107-GUACIMAL 704-Talamanca 11803-SANCHEZ 20304-SAN ROQUE 30106-GUADALUPE O ARENILLA 30602-CERVANTES 40202-SAN PEDRO 406-San Isidro 50205-SÁMARA 50806-TIERRAS MORENAS 60108-BARRANCA 70401-BRATSI 11801-CURRIDABAT 20305-TACARES 30107-CORRALILLO 30603-CAPELLADES 40203-SAN PABLO 40601-SAN ISIDRO 50207-BELÉN DE NOSARITA 50807-ARENAL 60109-MONTE VERDE 70404-TELIRE 107-Mora 20307-PUENTE DE PIEDRA 30108-TIERRA BLANCA 305-TURRIALBA 40204-SAN ROQUE 40602-SAN JOSÉ 503-Santa Cruz 509-Nandayure 60112-CHACARITA 10704-PIEDRAS NEGRAS 20308-BOLIVAR 30109-DULCE NOMBRE 30512-CHIRRIPO -
5Bcf43a74d065 202 Zonashom
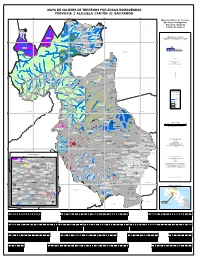
MAPA DE VALORES DE TERRENOS POR ZONAS HOMOGÉNEAS PROVINCIA 2 ALAJUELA CANTÓN 02 SAN RAMÓN 420000 430000 440000 450000 A La Fo rtu Finca Educativa Don Juanna Mapa de Valores de Terrenos El Jauuri Finca Ofelia o r Q r u u nm eb por Zonas Homogéneas B r ad o 2 02 13 R17/U17 a í P R i ed ra Cerro Blanco 2 02 13 U19 Empacadora Santa Fe 2 02 13 R04/U04ænm Provincia 2 Alajuela Volcano Gold 2 02 13 R18/U18 Cabinas Taboo nmæ 2 02 13 R06/U06 Caserío El Banco a rit ed Pi a Cantón 02 Alajuela ad ao Plaza br ac Canopy El Desafío e C Qu da ænm ra eb Qu a Concepción gu rrito ha Bu c Río ha Plaza C Castillo Nuevo Cerro Chato 2 02 13nmæ R05/U05 ío Ebaisnmæ R 2 02 nm13æ U03 s a c n a 2 02 13 R15/U15 l B Cementerio tinas Cris ebrada s Q Qu a u Q ñ e u æ e b a nm e P r n b a o u r Caserío Cerro Alegre Cairo 2 02 13 R07/U07 í d t a r a d æ R o a nm D F F a lo o n í r s e 2 02 13 R02/U02 t a R s j a o R Urb. La Cruz lle 2 02 13 U26 Q a u C e Linda Vista Ministerio de Hacienda b 2 02 13 R21/U21 ra d a 2 02 13 R16 Villa Vila æ P hachagüita a Quebrada C nm j nmColono a Finca La Bonanza Órgano de Normalización Técnica nm 2 02 13 R01/U01æ 2 02 13 U20 Q ue Cerro Los Perdidos br ad a 2 02 13 R08/U08 Gr æ an Hotel Bosques 2 02 13 R22/U22 de nm 2 02 13 R25/U25 1150000 Tilarán 1150000 ænm Hotel Chachagua Rain Forest Quintas La Pradera Coco Loco Canal 2 02 13 U09 Planta Hidroeléctrica Peñas Blancas 2 02 13 R14 Texas AyM University Plaza 2 02 13 R24/U24 nm 2 02 13 U11 Fila Delio Herrea ænm Cruz Roja Finca Cocoritos Plantel del ICE 2 02 13 U23 PEÑAS BLANCAS æ R ío C San Carlos a R 2 02 13 R10/U10 ñ ío o A 2 02 13 R12/U12 nm s N gu a o e l G l g a i r ta Ca La Altura o lle b Ra a d J io C o ima í nm R Q Q u e A ls u e a b L e b m E a b r T r a i a d g r d a a a S P a a l v i t a r s l a a l n Zona de Protección a C Aprobado por: Poco Sol Qu eb ra San Miguel A Florencia da T ar gu Pueblo Nuevo as al 2 02 14 R12/U12 nmæ Ing. -

Genetic Variability Among Populations of Lutzomyia
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(2): 189-196, February 2001 189 Genetic Variability among Populations of Lutzomyia (Psathyromyia) shannoni (Dyar 1929) (Diptera: Psychodidae: Phlebotominae) in Colombia Estrella Cárdenas/+, Leonard E Munstermann*, Orlando Martínez**, Darío Corredor**, Cristina Ferro Laboratorio de Entomología, Instituto Nacional de Salud, Avenida Eldorado, Carrera 50, Zona Postal 6, Apartado Aéreo 80080, Bogotá DC, Colombia *Department of Epidemiology and Public Health, School of Medicine, Yale University, New Haven, CT, USA **Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá DC, Colombia Polyacrylamide gel electrophoresis was used to elucidate genetic variation at 13 isozyme loci among forest populations of Lutzomyia shannoni from three widely separated locations in Colombia: Palambí (Nariño Department), Cimitarra (Santander Department) and Chinácota (Norte de Santander Depart- ment). These samples were compared with a laboratory colony originating from the Magdalena Valley in Central Colombia. The mean heterozygosity ranged from 16 to 22%, with 2.1 to 2.6 alleles detected per locus. Nei’s genetic distances among populations were low, ranging from 0.011 to 0.049. The esti- mated number of migrants (Nm=3.8) based on Wright’s F-Statistic, FST, indicated low levels of gene flow among Lu. shannoni forest populations. This low level of migration indicates that the spread of stomatitis virus occurs via infected host, not by infected insect. In the colony sample of 79 individuals, 0.62 0.62 the Gpi locus was homozygotic ( /0.62) in all females and heterozygotic ( /0.72) in all males. Al- though this phenomenon is probably a consequence of colonization, it indicates that Gpi is linked to a sex determining locus. -

Life Tables and Reproductive Parameters of Lutzomyia Spinicrassa (Diptera: Psychodidae) Under Laboratory Conditions
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99(6): 603-607, October 2004 603 Life Tables and Reproductive Parameters of Lutzomyia spinicrassa (Diptera: Psychodidae) under Laboratory Conditions Jesús Escovar, Felio J Bello/+, Alberto Morales, Ligia Moncada*, Estrella Cárdenas Laboratorio de Entomología, Biología Celular y Genética, Departamento de Ciencias Básicas, Universidad de La Salle, Bogotá DC, Colombia *Laboratorio de Parasitología, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá DC, Colombia Lutzomyia spinicrassa is a vector of Leishmania braziliensis in Colombia. This sand fly has a broad geographical distribution in Colombia and Venezuela and it is found mainly in coffee plantations. Baseline biological growth data of L. spinicrassa were obtained under experimental laboratory conditions. The development time from egg to adult ranged from 59 to 121 days, with 12.74 weeks in average. Based on cohorts of 100 females, horizontal life table was constructed. The following predictive parameters were obtained: net rate of reproduction (8.4 females per cohort female), generation time (12.74 weeks), intrinsic rate of population increase (0.17), and finite rate of population increment (1.18). The reproductive value for each class age of the cohort females was calculated. Vertical life tables were elaborated and mortality was described for the generation obtained of the field cohort. In addition, for two successive generations, additive variance and heritability for fecundity were estimated. Key words: Lutzomyia spinicrassa - life cycle - reproduction - population - heritability The sand fly Lutzomyia spinicrassa (Morales, Osor- shannoni, Cabrera et al. (1999) with L. ovallesi and Cabrera no-Mesa, Osorno & de Hoyos, 1969) belongs to the group and Ferro (2000) with three species of Lutzomyia of the verrucarum, series townsendi and it has a wide geographi- group verrucarum. -

Nombre Del Comercio Provincia Distrito Dirección Horario
Nombre del Provincia Distrito Dirección Horario comercio Almacén Agrícola Alajuela Aguas Claras Alajuela, Upala Aguas Claras, Cruce Del L-S 7:00am a 6:00 pm Aguas Claras Higuerón Camino A Rio Negro Comercial El Globo Alajuela Aguas Claras Alajuela, Upala Aguas Claras, contiguo L - S de 8:00 a.m. a 8:00 al Banco Nacional p.m. Librería Fox Alajuela Aguas Claras Alajuela, Upala Aguas Claras, frente al L - D de 7:00 a.m. a 8:00 Liceo Aguas Claras p.m. Librería Valverde Alajuela Aguas Claras Alajuela, Upala, Aguas Claras, 500 norte L-D de 7:00 am-8:30 pm de la Escuela Porfirio Ruiz Navarro Minisúper Asecabri Alajuela Aguas Claras Alajuela, Upala Aguas Claras, Las Brisas L - S de 7:00 a.m. a 6:00 400mts este del templo católico p.m. Minisúper Los Alajuela Aguas Claras Alajuela, Upala, Aguas Claras, Cuatro L-D de 6 am-8 pm Amigos Bocas diagonal a la Escuela Puro Verde Alajuela Aguas Claras Alajuela, Upala Aguas Claras, Porvenir L - D de 7:00 a.m. a 8:00 Supermercado 100mts sur del liceo rural El Porvenir p.m. (Upala) Súper Coco Alajuela Aguas Claras Alajuela, Upala, Aguas Claras, 300 mts L - S de 7:00 a.m. a 7:00 norte del Bar Atlántico p.m. MINISUPER RIO Alajuela AGUAS ALAJUELA, UPALA , AGUAS CLARAS, L-S DE 7:00AM A 5:00 PM NIÑO CLARAS CUATRO BOCAS 200M ESTE EL LICEO Abastecedor El Alajuela Aguas Zarcas Alajuela, Aguas Zarcas, 25mts norte del L - D de 8:00 a.m. -

Detection of Vesicular Stomatitis Virus Indiana from Insects Collected During the 2020 Outbreak in Kansas, USA
pathogens Article Detection of Vesicular Stomatitis Virus Indiana from Insects Collected during the 2020 Outbreak in Kansas, USA Bethany L. McGregor 1,† , Paula Rozo-Lopez 2,† , Travis M. Davis 1 and Barbara S. Drolet 1,* 1 Arthropod-Borne Animal Diseases Research Unit, Center for Grain and Animal Health Research, Agricultural Research Service, United States Department of Agriculture, Manhattan, KS 66502, USA; [email protected] (B.L.M.); [email protected] (T.M.D.) 2 Department of Entomology, Kansas State University, Manhattan, KS 66506, USA; [email protected] * Correspondence: [email protected] † These authors contributed equally to this work. Abstract: Vesicular stomatitis (VS) is a reportable viral disease which affects horses, cattle, and pigs in the Americas. Outbreaks of vesicular stomatitis virus New Jersey serotype (VSV-NJ) in the United States typically occur on a 5–10-year cycle, usually affecting western and southwestern states. In 2019–2020, an outbreak of VSV Indiana serotype (VSV-IN) extended eastward into the states of Kansas and Missouri for the first time in several decades, leading to 101 confirmed premises in Kansas and 37 confirmed premises in Missouri. In order to investigate which vector species contributed to the outbreak in Kansas, we conducted insect surveillance at two farms that experienced confirmed VSV-positive cases, one each in Riley County and Franklin County. Centers for Disease Control and Prevention miniature light traps were used to collect biting flies on the premises. Two genera of known VSV vectors, Culicoides biting midges and Simulium black flies, were identified to species, Citation: McGregor, B.L.; Rozo- pooled by species, sex, reproductive status, and collection site, and tested for the presence of VSV- Lopez, P.; Davis, T.M.; Drolet, B.S. -

Cantón Y Región Abogados Agrimensores Arquitectos Banqueros Ingenieros Boticarios Dentistas Médicos Otros Total Prof
Cuadro 110 COSTA RICA: PROFESIONALES POR CANTÓN Y REGIÓN SEGÚN EL CENSO DE 1883 Preceptores Cantón y Región Abogados Agrimensores Arquitectos Banqueros Ingenieros Boticarios Dentistas Médicos Otros Total Prof. maestros País 78 30 5 3 241 13 44 7 35 4 458 Liberia 1 11 612 1 13 Bagaces 2 2 Cañas 2 2 Abangares Tilarán La Cruz Guanacaste Este y Norte 1 11 1012 1 17 Carrillo Santa Cruz 13 1 14 Nicoya 12 12 Hojancha Nandayure Guanacaste Península 1 11 3513 1 26 Subtotal Guanacaste Este y Norte Lepanto, Paquera y Cóbano (L, P y C) Región Pacífico Norte 1 1 1 0 35 1 301 43 Puntarenas Central (sin L, P y C ni Pacífico Sur) 2 2 1 214 1 13 Esparza 2 1 3 Montes de Oro Puntarenas Norte 2 2 1 0 4 2 401016 Aguirre Parrita (ex Aguirre) Garabito San Mateo/Orotina 1 6 1 8 Turrubares (ex Puriscal) Región Pacífico Central 2 2 2 0 10 2 501 24 Buenos Aires Osa/Golfo Dulce 1 Golfito Corredores (ex Golfito) Coto Brus Perez Zeledón/El General Región Pacífico Sur 0 0 0 0 0 0 1 0 0 1 Central Alajuela, Sarapiquí S. Ramón Ángeles, Zapotal, Peñas Blancas Grecia, Río Cuarto Upala (ex S. Gerónimo Grecia) Los Chiles (ex S. Gerónimo Grecia) Guatuso (ex S. Gerónimo Grecia) San Carlos (ex S. Gertrudis Grecia)/Naranjo(S. Carlos) Alfaro Ruíz (Tapezco, Zapote, Laguna, Palmira) Valverde Vega, Toro Amarillo Central Heredia, Sarapiquí Región Norte/Huetar Norte 0 0 0 0 0 0 0 0 0 0 Limón Central 5 1 6 Matina Talamanca Pococí Guácimo Siquirres Centro, Matina y Talamanca Pococí, Guácimo, Siquirres Región Atlántico/Huetar Atlántico 0 0 0 0 0 5 0 0 1 6 Puriscal/incluye Mercedes/S. -

Código Presupuestario Centro Educativo Dirección
CÓDIGO CENTRO EDUCATIVO DIRECCIÓN REGIONAL CIRCUITO PRESUPUESTARIO 1247 PATRIARCA SAN JOSÉ OCCIDENTE 01 1261 PATA DE GALLO OCCIDENTE 01 1262 ABRAHAM PANIAGUA NÚÑEZ OCCIDENTE 01 1292 GEORGINA BOLMARCICH DE ORLICH OCCIDENTE 01 1297 JORGE WASHINGTON OCCIDENTE 01 1298 J.N. FELICITAS RAMÍREZ VEGA OCCIDENTE 01 1316 JOSÉ JOAQUÍN SALAS PÉREZ OCCIDENTE 01 1320 LLANO BRENES OCCIDENTE 01 1327 MACARIO VALVERDE MADRIGAL OCCIDENTE 01 1333 LA UNIÓN OCCIDENTE 01 1335 MONSEÑOR CLODOVEO HIDALGO SOLANO OCCIDENTE 01 1350 RINCÓN DE MORA OCCIDENTE 01 1352 RÍO GRANDE OCCIDENTE 01 1355 MIXTA SAN RAFAEL OCCIDENTE 01 1369 LABORATORIO OCCIDENTE 01 1371 RINCÓN DE OROZCO OCCIDENTE 01 4033 INSTITUTO JULIO ACOSTA GARCÍA OCCIDENTE 01 4037 LICEO PATRIARCA SAN JOSÉ OCCIDENTE 01 ENSEÑANZA ESPECIAL Y REHABILITACION DE SAN 4495 OCCIDENTE 01 RAMÓN 4849 NOCTURNO JULIAN VOLIO LLORENTE OCCIDENTE 01 5643 J.N. JOSÉ JOAQUÍN SALAS PÉREZ OCCIDENTE 01 5996 EXPERIMENTAL BILINGÜE DE SAN RAMON OCCIDENTE 01 6250 CNV. ESCUELA JORGE WASHINGTON OCCIDENTE 01 6535 C.T.P. CALLE ZAMORA OCCIDENTE 01 6535 SECCION NOCTURNA C.T.P. CALLE ZAMORA OCCIDENTE 01 1242 ALBERTO MANUEL BRENES MORA OCCIDENTE 02 1252 ÁNGELES NORTE OCCIDENTE 02 1253 FÉLIX ÁNGEL SALAS CABEZAS OCCIDENTE 02 1281 LOS JARDINES OCCIDENTE 02 1282 LOS LAGOS OCCIDENTE 02 1289 FEDERICO SALAS CARVAJAL OCCIDENTE 02 1291 FRANCISCO JOSE ORLICH BOLMARCICH OCCIDENTE 02 1304 LA BALSA OCCIDENTE 02 1308 JULIÁN VOLIO LLORENTE OCCIDENTE 02 1312 LA PAZ OCCIDENTE 02 1318 LISÍMACO CHAVARRÍA PALMA OCCIDENTE 02 1330 MANUEL QUESADA BASTOS OCCIDENTE 02 1331 MERCEDES QUESADA QUESADA OCCIDENTE 02 1337 NAUTILIO ACOSTA PIEPPER OCCIDENTE 02 1338 RUPERTO ZUÑIGA SANCHO OCCIDENTE 02 1343 JUAN JOSÉ VALVERDE MADRIGAL OCCIDENTE 02 1361 SIMÓN BOLÍVAR OCCIDENTE 02 4032 LICEO NUESTRA SEÑORA DE LOS ANGELES OCCIDENTE 02 5548 DULCE NOMBRE OCCIDENTE 02 5808 J.N. -

Régimen De Zonas Francas En Costa Rica Base Legal
Base legal Régimen de Zonas Francas en Costa Rica Base legal Ley del Régimen de Zonas Francas y su Reglamento Ley de Creación de COMEX y PROCOMER Ley General de Aduanas y su Reglamento Leyes Se producen las últimas reformas a la Ley No. 7210 para cumplir con Ley No. 8794 las disposiciones de la OMC en materia de subvenciones y medidas (2010) compensatorias (1996) Ley No. 7830 Se reforma nuevamente la Ley No. 7210 (1998) Se fusiona CENPRO, la Corporación y el Ley No. 7638 Consejo Nacional de Inversiones, para dar vida a (1996) PROCOMER Ley No. 7210 Se crea la Ley del Régimen de Zonas (1990) Francas que conocemos hoy día Ley No. 6951 La ley sufre reformas (1984) Se crea la Ley No. 6695 Corporación de (1981) Zonas Francas Reglamentos 40141-COMEX-H (2017) 36725-COMEX-H (2011) Del 2001 a la fecha, el 36000-COMEX-H (2010) Reglamento a la Ley del 35422-COMEX-H (2009) Régimen de Zonas 34739-COMEX-H (2008) Francas ha sufrido 34166-COMEX-H (2007) varios cambios 33283-COMEX-H (2006) 33360-COMEX-H (2006) 31715-COMEX-H (2004) 31585-COMEX-H (2003) 30849-COMEX-H (2002) 29606-COMEX-H (2001) 29962-COMEX-H (2001) Se 28451-COMEX-H reglamenta la (1999) Ley No. 7210 Conceptos importantes Régimen de Zonas Francas en Costa Rica PROCOMER • Entidad pública de carácter no estatal. • Apoya técnica y financieramente al Ministerio de Comercio Exterior de Costa Rica (COMEX). • Promueve las exportaciones y los encadenamientos productivos, desarrolla estudios de mercado, y administra los regímenes especiales como el Régimen de Zonas Francas. -

Inec Requiere 2.200 Personas Para Realizar Censo Nacional Agropecuario
INEC REQUIERE 2.200 PERSONAS PARA REALIZAR CENSO NACIONAL AGROPECUARIO Personal se contratará por un mes para censar las fincas del país. El Instituto Nacional de Estadística y Censos, INEC, se encuentra en este momento reclutando a 2.200 personas para que trabajen como personal supervisor y entrevistador en el VI Censo Nacional Agropecuario a desarrollarse del 2 al 30 de junio próximo. El INEC considera importante que las personas que participen tengan alguna experiencia en el sector agrícola o ganadero y que conozcan la zona donde viven pues es donde podrían trabajar. El INEC hace un llamado especial a aquellas personas que cuenten con un título preferiblemente de un Colegio Técnico Agropecuario o bien estudiantes o egresados de carreras afines a la agricultura y ganadería, para que se incorporen al equipo de trabajo de campo CENAGRO. Para la contratación de este personal se pide de forma opcional contar con teléfono inteligente y algún medio de transporte. Los requerimientos se pueden flexibilizar de acuerdo a las regiones geográficas donde hay menor disponibilidad de personal. Hasta la fecha el INEC ha recibido una cantidad importante de inscripciones, sin embargo hay zonas donde se percibe un faltante (recuadro) y se hace un llamado a la población de los siguientes lugares para que se inscriba y llene esas vacantes. LUGARES DONDE PRINCIPALMENTE SE REQUIERE PERSONAL REGION CANTON DISTRITO Pacífico central San Mateo Desmonte, Labrador Aguirre Naranjillo Puntarenas Manzanillo, Arancibia Chorotega Abangares Sierra Cañas Bebedero Hojancha -

Oficina Subregional De Grecia
Instituto de Desarrollo Rural de Costa Rica Oficina Subregional de Grecia Informe de Caracterización Básica Territorio Atenas-Palmares-Naranjo-San Ramón-Zarcero Costa Rica, 2016 Tabla de contenidos Lista de Cuadros ................................................................................................................................... i Lista de Ilustraciones ...........................................................................................................................iii Lista de Gráficos ................................................................................................................................. iv Glosario de acrónimos y siglas............................................................................................................ v Resumen .............................................................................................................................................. vi Introducción ........................................................................................................................................ 1 1. Contexto de los territorios Inder .................................................................................................. 5 1.1. Los Territorios Rurales ............................................................................................... 5 1.2. Caracterización del área de trabajo ........................................................................... 8 2. Objetivos .................................................................................................................................. -

Insect Egg Size and Shape Evolve with Ecology but Not Developmental Rate Samuel H
ARTICLE https://doi.org/10.1038/s41586-019-1302-4 Insect egg size and shape evolve with ecology but not developmental rate Samuel H. Church1,4*, Seth Donoughe1,3,4, Bruno A. S. de Medeiros1 & Cassandra G. Extavour1,2* Over the course of evolution, organism size has diversified markedly. Changes in size are thought to have occurred because of developmental, morphological and/or ecological pressures. To perform phylogenetic tests of the potential effects of these pressures, here we generated a dataset of more than ten thousand descriptions of insect eggs, and combined these with genetic and life-history datasets. We show that, across eight orders of magnitude of variation in egg volume, the relationship between size and shape itself evolves, such that previously predicted global patterns of scaling do not adequately explain the diversity in egg shapes. We show that egg size is not correlated with developmental rate and that, for many insects, egg size is not correlated with adult body size. Instead, we find that the evolution of parasitoidism and aquatic oviposition help to explain the diversification in the size and shape of insect eggs. Our study suggests that where eggs are laid, rather than universal allometric constants, underlies the evolution of insect egg size and shape. Size is a fundamental factor in many biological processes. The size of an 526 families and every currently described extant hexapod order24 organism may affect interactions both with other organisms and with (Fig. 1a and Supplementary Fig. 1). We combined this dataset with the environment1,2, it scales with features of morphology and physi- backbone hexapod phylogenies25,26 that we enriched to include taxa ology3, and larger animals often have higher fitness4.