Developmental Biology and Morphogenesis of the Face, Lip And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
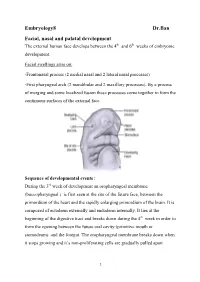
Embryology8 Dr.Ban Facial, Nasal and Palatal Development the External Human Face Develops Between the 4Th and 6Th Weeks of Embryonic Development
Embryology8 Dr.Ban Facial, nasal and palatal development The external human face develops between the 4th and 6th weeks of embryonic development. Facial swellings arise on: -Frontonasal process (2 medial nasal and 2 lateral nasal processes) -First pharyngeal arch (2 mandibular and 2 maxillary processes). By a process of merging and some localized fusion these processes come together to form the continuous surfaces of the external face. Sequence of developmental events : During the 3rd week of development an oropharyngeal membrane (buccopharyngeal ) is first seen at the site of the future face, between the primordium of the heart and the rapidly enlarging primordium of the brain. It is composed of ectoderm externally and endoderm internally. It lies at the beginning of the digestive tract and breaks down during the 4th week in order to form the opening between the future oral cavity (primitive mouth or stomodeum) and the foregut. The oropharyngeal membrane breaks down when it stops growing and it’s non-proliferating cells are gradually pulled apart 1 because they cannot fill the expanding area.The tissues around it expand very rapidly. The face develops from five primordia that appear in the fourth week: the frontonasal prominence, the two maxillary swellings, and the two mandibular swellings. The external face forms from two sources that surround the oropharyngeal membrane 1-Tissues of the frontonasal process that cover the forebrain, predominantly of neural crest origin. 2-The tissues of the first (or mandibular) pharyngeal arch, of mixed mesoderm and neural crest origin Face initially formed by 5 mesenchymal swellings ( prominences): Two mandibular prominences Two maxillary prominences Frontonasal prominence (midline structure is a single structure that is ventral to the forebrain. -

Palatal Fibroma - a Case-Report Farooque Khan, Romita Gaikwad Vspms Dental College & Research Centre, Nagpur, India Correspondence ABSTRACT Dr
Case Report Journal of College of Medical Sciences-Nepal, Vol-12, No 1, Jan-Mar 016 ISSN: 2091-0657 (Print); 2091-0673 (Online) Open Access Palatal Fibroma - A Case-report Farooque Khan, Romita Gaikwad VSPMs Dental College & Research Centre, Nagpur, India Correspondence ABSTRACT Dr. Farooque Khan Fibroma is a benign tumor of fibrous connective tissue. Fibromas represent M.D.S. Periodontics, VSPMs inflammatory state rather than neoplastic conditions, which are mostly Dental College & Research sessile or slightly pendunculated with a smooth contour, pale pink and are Centre, firm in consistency, which commonly occurs on gingiva, tongue, buccal Nagpur - 440019. India mucosa and palate. Cinical, radiographical as well as histologic findings in Email: [email protected] combination with surgical findings are beneficial, but it further requires more studies to determine the nature of such fibromatous lesions. A DOI: http:// interdisciplinary access is thus needed in treatment of fibrous lesions , so dx.doi.org/10.3126/ as to reduce its reocurrence and to boost the standard of life, thus providing jcmsn.v12i1.14417 better functioning and esthetics. Key words: Benign tumor, cemento-ossifying, fibroma, palate Citation: Khan F, Gaikwad R. Palatal Fibroma - A Case-report. JCMS Nepal. 2016;12(1):36-9. INTRODUCTION having a habit of tobacco chewing 4-5 times a day Benign tumors of fibrous connective tissue are since 10 years. commonly seen in the oral cavity. Fibroma is a Intraoral examination : benign tumor of fibrous connective tissue.1 A A single growth of approx size 0.5 X 0.5cm was majority of fibromas occurring in the oral cavity are present on left side of hard palate in 24, 25 region. -

Musculus Uvulae and Palatine Raphe
The Longitudinal Fibromuscular Component of the Soft Palate in the Fifteen-Week Human Fetus: Musculus Uvulae and Palatine Raphe HERBERT L. LANGDON, Ph.D. KATHLEEN KLUEBER, M.S. Pittsburgh, Pennsylvania 15261 The structural relationships of the longitudinal fibromuscular component of the soft palate (musculus uvulae and raphe) were studied using histologic sections from 19 early human fetal specimens. Musculus uvulae arises in association with the palatine aponeurosis near the beginning of the second quadrant of the velum, follows a sigmoid course, and terminates near the base of the uvula. In addition, an occasional muscular loop may arise from the bony palate, arch downwards, and then recur into the uvular muscle. A complex relationship exists between the raphe in the velum and several palatal muscles. With regard to musculus uvulae, small muscular bundles arise from the raphe to embrace the muscle near its crest. These branches may aid in contouring the dorsal surface of the velum in the region of the levator eminence to complement the surface of the posterior pharyngeal wall and thus enhance the efficiency of the velopharyngeal seal. Introduction received considerable attention in the litera- ture from the perspectives of comparative In the early human fetus, as well as in the anatomy (Kuenzel et al., 1966), prenatal de- adult, musculus uvulae and the palatine velopment (Doménech-Ratto, 1977; Peter, raphe form a longitudinally-oriented fibro- 1913; Futamura, 1906), and the gross anat- muscular complex running the length of the omy of the muscle in the adult (Azzam and soft palate. The uvular muscle overrides the Kuehn, 1977; Kriens, 1975; Voth, 1961; Rue- other palatal musculature in the dorsal mid- dinger, 1879; Henle, 1873). -

Anatomical Basis of Craniofacial Birth Defects
AnatomicalAnatomical BasisBasis ofof CraniofacialCraniofacial BirthBirth DefectsDefects Handout download: http://www.oucom.ohiou.edu/dbms-witmer/peds-rpac.htm LawrenceLawrence M.M. Witmer,Witmer, PhDPhD Department of Biomedical Sciences College of Osteopathic Medicine Ohio University Athens, Ohio 45701 [email protected] 18 October 2000 DevelopmentDevelopment ofof thethe FaceFace II • 5 facial primordia • Frontonasal prominence • Paired maxillary prominences • Paired mandibular prominences • Surround primordial mouth (stomodeum) • Neural crest: source for almost all connective tissues in the face • Frontonasal prominence forms forehead and nose and a short margin of mouth • Lower jaw and lip form first • Nasal placodes (and pit): surrounded by medial & lateral nasal prominences • Nasal pit remains connected to mouth • Maxillary prominences grow toward each other, pushing nasal prominences medially From Moore 1982 DevelopmentDevelopment ofof thethe FaceFace IIII • Medial nasal prominences merge with each other and with lateral nasal & maxillary prominences • Nasolacrimal groove: between lateral nasal and maxillary prominences • Becomes nasolacrimal duct • Duct forms as solid epithelial cord that later canalizes • Nasolacrimal duct atresia • Failure to completely canalize • 6% of newborns • Intermaxillary segment • Merger of medial nasal prominences • Gives rise to philtrum, premaxillary bones, primary palate From Moore 1982 SummarySummary ofof FacialFacial DevelopmentDevelopment From Moore 1982 Disruptions in the formation -

Trigeminal Nerve, Mandibular Division Basic Anatomy and a Bit More
The palate and the faucial isthmus He who guards his mouth and his tongue keeps himself from calamity. Proverbs 21:23 Ph.D., Dr. David Lendvai Parts of the oral cavity Parts of the oral cavity 1. Vestibule of the oral cavity Borders: - lips and cheek (bucca) - dental arches 2. Oral cavity proper Borders: - roof: hard and soft palate - floor: oral diaphragm (mylohoid m.) - antero-laterally: dental arches - posteriorly: isthmus of the fauces Etrance of the oral cavity - Philtrum - Upper & lower lip - Angulus - Rubor labii - Nasolabial groove (Facial palsy) Sobotta Szentágothai - Réthelyi Aspectus anterior 1 zygomatic process 2 frontal process 2 4 alveolar process 1 4 Faller (left) lateral aspect 1 zygomatic process 2 frontal process 3 orbital surface 4 alveolar process 2 3 Sobotta 1 4 Faller (right) Medial aspect Sobotta Superior aspect Sobotta Inferior aspect Sobotta http://www.almanahmedical.eu Sobotta Florian Dental – Dr. S. Kovách Fehér Fehér Szél Szél http://www.hc-bios.com Structures of the hard palate - incisive papilla - palatine rugae - palatine raphe - torus Hard and soft palate Muscles of the soft palate - Levator veli palatini m. - Tensor veli palatini m. - Palatoglossus m. - Palatopharyngeus m. - M. uvulae Muscles of the soft palate Muscles of the soft palate Structures of the hard and soft palate - mucous membrane - palatine glands - bone / muscles Histology of the hard palate Mucoperiosteum Histology of the soft palate NASAL SURFACE - pseudostratified ciliated columnar epithelium - lamina propria - mucous glands - striated -

Development and Malformations of Face and Palate
Development and malformations of face and palate Compiled by András Csillag and Andrea D. Székely GERMINAL LAYER DERIVATIVES ECTODERM contributing to the formation of the face appears by the 4th week. The oropharyngeal membrane (interface between ECTODERM and ENDODERM) is located in front of the later palatine tonsils. Ectodermal structures limiting the stomodeum participate in the formation of the face, as well as of the nasal and oral cavities. MESENCHYME that fills the pharyngeal arches derives from the neural crest ECTOMESENCHYME DEVELOPMENT OF THE FACE WEEK 6 The face is formed by 5 processes approximately 24 days Frontonasal prominence (1) Maxillary prominence (2) 1st arch Mandibular prominence (2) nasal (olfactory) pits form surrounded by the medial and lateral nasal processes nasolacrimal groove separates the lateral nasal process from the maxillary process maxillary processes fuse with the medial nasal processes lateral nasal processes fuse with the maxillary processes, thus obliterating the nasolacrimal groove. DEVELOPMENT OF THE FACE 5-week embryo 6-week embryo 7-week embryo. 10-week embryo The nasal prominences are gradually Maxillary prominences have fused separated from the maxillary with the medial nasal prominences. prominence by deep furrows. CEPHALIC PRIMORDIA - PLACODES Olfactory placode Pharyngeal Stomodeum” arches Nasal placodes - thickenings of the surface ectoderm (later differentiate into the olfactory epithelium) DEVELOPMENT OF THE NASAL CAVITIES AND THE HARD PALATE By week 5 the placodes form the nasal pits. They further invaginate and the pits approach the primitive oral cavity. A thin oronasal membrane separates the two cavities. By its rupture the primitive choanae will form DEVELOPMENT OF THE NASAL CAVITIES AND THE HARD PALATE By week 8, a partition forms between the primitive nasal chambers and the oral cavity The primary palate - the anterior aspect derives from the intermaxillary segment (or median palatine process, formed by the medial nasal processes). -

ORAL MUCOSA with Particular Reference to The
ORAL MUCOSA With Particular Reference to the Edentulous Mouth Volume 1 of Two Volumes IAN BUCHANAN WATSON - B.D.S., F.D.S.R.C.P.S. (Glasg.) THESIS Submitted for the Degree of Doctor of Philosophy Glasgow Dental Hospital and School University of Glasgow, October 1978. ProQuest Number: 13804170 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a com plete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. uest ProQuest 13804170 Published by ProQuest LLC(2018). Copyright of the Dissertation is held by the Author. All rights reserved. This work is protected against unauthorized copying under Title 17, United States C ode Microform Edition © ProQuest LLC. ProQuest LLC. 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, Ml 48106- 1346 1 CONTENTS VOLUME 1 PAGE CHAPTER CONTENTS 1 ACKNOWLEDGEMENTS 11 PREFACE 13 SUMMARY CHAPTER 1 ORAL MUCOSA AND DENTURES 16 CHAPTER 2 PRELIMINARY POST-MORTEM STUDY 80 CHAPTER 3 POST-MORTEM STUDY OF INTACT PALATES 101 CHAPTER 4 NORMAL HUMAN PALATAL MUCOSA 123 CHAPTER 5 PALATAL MUCOSA UNDER COMPLETE DENTURES 1U5 CHAPTER 6 THE EFFECTS OF COMPLETE DENTURES ON ORAL MUCOSA 158 CHAPTER 7 DISCUSSION 167 ABBREVIATIONS 178 REFERENCES 180 VOLUME 2 FIGURES AND TABLES ARRANGED IN SEQUENCE AS THEY ARE REFERRED TO IN VOLUME 1. 2 CHAPTER ONE CONTENTS - ORAL MUCOSA AND DENTURES PAGE 1.1 INTRODUCTION 16 1.2 NORMAL ORAL -

A Combined Approach of Teaching Head Development Using Embryology and Comparative Anatomy
Edorium J Anat Embryo 2016;3:17–27. Danowitz et al. 17 www.edoriumjournals.com/ej/ae REVIEW ARTICLE PEER REVIEWED | OPEN ACCESS A combined approach of teaching head development using embryology and comparative anatomy Melinda Danowitz, Hong Zheng, Adriana Guigova, Nikos Solounias ABSTRACT the evolutionary changes of many structures allows for a greater understanding of the Many aspects of human head embryology reflect human embryology, and removes the need for its evolutionary development. The pharyngeal memorization of seemingly complex processes. arches, a major component of head development, A link to comparative evolutionary anatomy originally functioned in filter feeding and provides context to the purpose and morphology vascular exchange, which is why each arch of primitive structures, and clarifies several has associated vasculature and muscles. The issues in human head development. primitive tongue had few-associated muscles and was responsible for simple movements; the Keywords: Anatomy education, Embryology, Head human tongue evolved post-otic somites that and neck, Pharyngeal arches migrate to the tongue and develop the majority of the tongue musculature. These somites originate How to cite this article outside the tongue, and the motor innervation therefore differs from the general and special Danowitz M, Zheng H, Guigova A, Solounias N. A sensory innervation. In the primitive condition, combined approach of teaching head development the trapezius and sternocleidomastoid belonged using embryology and comparative anatomy. to a single muscle group that were involved in Edorium J Anat Embryo 2016;3:17–27. gill movements; they separate into two muscles with the reduction of certain skeletal elements, but retain the same innervation. -

Syndromes of the First and Second Branchial Arches, Part 1: Embryology and Characteristic REVIEW ARTICLE Defects
Syndromes of the First and Second Branchial Arches, Part 1: Embryology and Characteristic REVIEW ARTICLE Defects J.M. Johnson SUMMARY: A variety of congenital syndromes affecting the face occur due to defects involving the G. Moonis first and second BAs. Radiographic evaluation of craniofacial deformities is necessary to define aberrant anatomy, plan surgical procedures, and evaluate the effects of craniofacial growth and G.E. Green surgical reconstructions. High-resolution CT has proved vital in determining the nature and extent of R. Carmody these syndromes. The radiologic evaluation of syndromes of the first and second BAs should begin H.N. Burbank first by studying a series of isolated defects: CL with or without CP, micrognathia, and EAC atresia, which compose the major features of these syndromes and allow more specific diagnosis. After discussion of these defects and the associated embryology, we proceed to discuss the VCFS, PRS, ACS, TCS, Stickler syndrome, and HFM. ABBREVIATIONS: ACS ϭ auriculocondylar syndrome; BA ϭ branchial arch; CL ϭ cleft lip; CL/P ϭ cleft lip/palate; CP ϭ cleft palate; EAC ϭ external auditory canal; HFM ϭ hemifacial microsomia; MDCT ϭ multidetector CT; PRS ϭ Pierre Robin sequence; TCS ϭ Treacher Collins syndrome; VCFS ϭ velocardiofacial syndrome adiographic evaluation of craniofacial deformities is nec- major features of the syndromes of the first and second BAs. Ressary to define aberrant anatomy, plan surgical proce- Part 2 of this review discusses the syndromes and their radio- dures, and evaluate the effects of craniofacial growth and sur- graphic features: PRS, HFM, ACS, TCS, Stickler syndrome, gical reconstructions.1 The recent rapid proliferation of and VCFS. -

Illustrated Review of the Embryology and Development of the Facial
REVIEW ARTICLE Illustrated Review of the Embryology and Development of the Facial Region, Part 2: Late Development of the Fetal Face and Changes in the Face from the Newborn to Adulthood P.M. Som and T.P. Naidich ABSTRACT SUMMARY: The later embryogenesis of the fetal face and the alteration in the facial structure from birth to adulthood have been reviewed. Part 3 of the review will address the molecular mechanisms that are responsible for the changes described in parts 1 and 2. art 1 of this 3-part review primarily dealt with the early em- first make contact, each is completely covered by a homoge- Pbryologic development of the face and nasal cavity. Part 2 will neous epithelium. A special epithelium arises at the edge of discuss the later embryonic and fetal development of the face, and each palatal shelf, facilitating the eventual fusion of these changes in facial appearance from neonate to adulthood will be shelves. The epithelium on the nasal cavity surface of the palate reviewed. will differentiate into columnar ciliated epithelium. The epi- thelium on the oral cavity side of the palate will differentiate Formation of the Palate into stratified squamous epithelium. Between the sixth and 12th weeks, the palate is formed from 3 The 2 palatal shelves also fuse with the triangular primary pal- primordia: a midline median palatine process and paired lateral ate anteromedially to form a y-shaped fusion line. The point of palatine processes (Fig 1). In the beginning of the sixth week, fusion of the secondary palatal shelves with the primary palate is merging of the paired medial nasal processes forms the intermax- marked in the adult by the incisive foramen. -

Dental Assisting Science I
ADED 110C: Dental Assisting Science I Hours: Lecture - 3, Lab - 0, Credits - 3 Prerequisite: none Term & Dates: Fall 2020 Faculty: Kelly O’Brien, CDA, RDH, MEd Faculty Accessibility: available via discussion board and email, virtual conferences by appointment Email: [email protected] DRAFT Course Outline – official syllabus will be available in August Course Description A study of the anatomy of the head, emphasizing the osteological landmarks and the structures of the oral cavity. Both the permanent and primary dentitions are studied, including embryonic development and eruption patterns. In addition, an introduction to the structure and function of the human body systems in health and disease will be presented. Return to Top Learning Outcomes Educated Person Statement of Philosophy Upon completion of this course, students will be able to: Pronounce, spell, and define key terminology required to function within the profession. Identify any tooth on the oral cavity by name, number, location, function and shorthand terms. Discuss the development of the head and neck from conception through adulthood with a focus on the oral cavity. Describe the major systems of the body, their functions, and relationship to oral health. Acquired Knowledge and Skills: List the classifications and function of each of the teeth. Identify the different tissues of the teeth and oral cavity. Identify the arrangement of the dentitions by arch, quadrant, and sextant. Label the surfaces of any tooth, and the divisions into thirds of the root and the crown. 1 Describe the dentitions using eruption and shedding dates. Provide the shorthand identification of each tooth using Palmer, FDI, and Universal. -

Aandp2ch25lecture.Pdf
Chapter 25 Lecture Outline See separate PowerPoint slides for all figures and tables pre- inserted into PowerPoint without notes. Copyright © McGraw-Hill Education. Permission required for reproduction or display. 1 Introduction • Most nutrients we eat cannot be used in existing form – Must be broken down into smaller components before body can make use of them • Digestive system—acts as a disassembly line – To break down nutrients into forms that can be used by the body – To absorb them so they can be distributed to the tissues • Gastroenterology—the study of the digestive tract and the diagnosis and treatment of its disorders 25-2 General Anatomy and Digestive Processes • Expected Learning Outcomes – List the functions and major physiological processes of the digestive system. – Distinguish between mechanical and chemical digestion. – Describe the basic chemical process underlying all chemical digestion, and name the major substrates and products of this process. 25-3 General Anatomy and Digestive Processes (Continued) – List the regions of the digestive tract and the accessory organs of the digestive system. – Identify the layers of the digestive tract and describe its relationship to the peritoneum. – Describe the general neural and chemical controls over digestive function. 25-4 Digestive Function • Digestive system—organ system that processes food, extracts nutrients, and eliminates residue • Five stages of digestion – Ingestion: selective intake of food – Digestion: mechanical and chemical breakdown of food into a form usable by