Phylogenetic Diversity and Endemism: Metrics for Identifying Critical Regions of Conifer Conservation in Australia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Extinction, Transoceanic Dispersal, Adaptation and Rediversification
Turnover of southern cypresses in the post-Gondwanan world: Title extinction, transoceanic dispersal, adaptation and rediversification Crisp, Michael D.; Cook, Lyn G.; Bowman, David M. J. S.; Author(s) Cosgrove, Meredith; Isagi, Yuji; Sakaguchi, Shota Citation The New phytologist (2019), 221(4): 2308-2319 Issue Date 2019-03 URL http://hdl.handle.net/2433/244041 © 2018 The Authors. New Phytologist © 2018 New Phytologist Trust; This is an open access article under the terms Right of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited. Type Journal Article Textversion publisher Kyoto University Research Turnover of southern cypresses in the post-Gondwanan world: extinction, transoceanic dispersal, adaptation and rediversification Michael D. Crisp1 , Lyn G. Cook2 , David M. J. S. Bowman3 , Meredith Cosgrove1, Yuji Isagi4 and Shota Sakaguchi5 1Research School of Biology, The Australian National University, RN Robertson Building, 46 Sullivans Creek Road, Acton (Canberra), ACT 2601, Australia; 2School of Biological Sciences, The University of Queensland, Brisbane, Qld 4072, Australia; 3School of Natural Sciences, The University of Tasmania, Private Bag 55, Hobart, Tas 7001, Australia; 4Graduate School of Agriculture, Kyoto University, Kyoto 606-8502, Japan; 5Graduate School of Human and Environmental Studies, Kyoto University, Kyoto 606-8501, Japan Summary Author for correspondence: Cupressaceae subfamily Callitroideae has been an important exemplar for vicariance bio- Michael D. Crisp geography, but its history is more than just disjunctions resulting from continental drift. We Tel: +61 2 6125 2882 combine fossil and molecular data to better assess its extinction and, sometimes, rediversifica- Email: [email protected] tion after past global change. -

Edition 2 from Forest to Fjaeldmark the Vegetation Communities Highland Treeless Vegetation
Edition 2 From Forest to Fjaeldmark The Vegetation Communities Highland treeless vegetation Richea scoparia Edition 2 From Forest to Fjaeldmark 1 Highland treeless vegetation Community (Code) Page Alpine coniferous heathland (HCH) 4 Cushion moorland (HCM) 6 Eastern alpine heathland (HHE) 8 Eastern alpine sedgeland (HSE) 10 Eastern alpine vegetation (undifferentiated) (HUE) 12 Western alpine heathland (HHW) 13 Western alpine sedgeland/herbland (HSW) 15 General description Rainforest and related scrub, Dry eucalypt forest and woodland, Scrub, heathland and coastal complexes. Highland treeless vegetation communities occur Likewise, some non-forest communities with wide within the alpine zone where the growth of trees is environmental amplitudes, such as wetlands, may be impeded by climatic factors. The altitude above found in alpine areas. which trees cannot survive varies between approximately 700 m in the south-west to over The boundaries between alpine vegetation communities are usually well defined, but 1 400 m in the north-east highlands; its exact location depends on a number of factors. In many communities may occur in a tight mosaic. In these parts of Tasmania the boundary is not well defined. situations, mapping community boundaries at Sometimes tree lines are inverted due to exposure 1:25 000 may not be feasible. This is particularly the or frost hollows. problem in the eastern highlands; the class Eastern alpine vegetation (undifferentiated) (HUE) is used in There are seven specific highland heathland, those areas where remote sensing does not provide sedgeland and moorland mapping communities, sufficient resolution. including one undifferentiated class. Other highland treeless vegetation such as grasslands, herbfields, A minor revision in 2017 added information on the grassy sedgelands and wetlands are described in occurrence of peatland pool complexes, and other sections. -

Plant Life of Western Australia
INTRODUCTION The characteristic features of the vegetation of Australia I. General Physiography At present the animals and plants of Australia are isolated from the rest of the world, except by way of the Torres Straits to New Guinea and southeast Asia. Even here adverse climatic conditions restrict or make it impossible for migration. Over a long period this isolation has meant that even what was common to the floras of the southern Asiatic Archipelago and Australia has become restricted to small areas. This resulted in an ever increasing divergence. As a consequence, Australia is a true island continent, with its own peculiar flora and fauna. As in southern Africa, Australia is largely an extensive plateau, although at a lower elevation. As in Africa too, the plateau increases gradually in height towards the east, culminating in a high ridge from which the land then drops steeply to a narrow coastal plain crossed by short rivers. On the west coast the plateau is only 00-00 m in height but there is usually an abrupt descent to the narrow coastal region. The plateau drops towards the center, and the major rivers flow into this depression. Fed from the high eastern margin of the plateau, these rivers run through low rainfall areas to the sea. While the tropical northern region is characterized by a wet summer and dry win- ter, the actual amount of rain is determined by additional factors. On the mountainous east coast the rainfall is high, while it diminishes with surprising rapidity towards the interior. Thus in New South Wales, the yearly rainfall at the edge of the plateau and the adjacent coast often reaches over 100 cm. -

1999 New Zealand Botanical Society
NEW ZEALAND BOTANICAL SOCIETY NEWSLETTER NUMBER 57 SEPTEMBER 1999 New Zealand Botanical Society President: Jessica Beever Secretary/Treasurer: Anthony Wright Committee: Bruce Clarkson, Colin Webb, Carol West Address: c/- Canterbury Museum Rolleston Avenue CHRISTCHURCH 8001 NEW ZEALAND Subscriptions The I999 ordinary and institutional subs are $18 (reduced to $15 if paid by the due date on the subscription invoice). The 1999 student sub, available to full-time students, is $9 (reduced to $7 if paid by the due date on the subscription invoice). Back issues of the Newsletter are available at $2.50 each from Number 1 (August 1985) to Number 46 (December 1996), $3.00 each from Number 47 (March 1997) to Number 50 (December 1997), and $3.75 each from Number 51 (March 1998) onwards. Since 1986 the Newsletter has appeared quarterly in March, June, September and December. New subscriptions are always welcome and these, together with back issue orders, should be sent to the Secretary/Treasurer (address above). Subscriptions are due by 28 February of each year for that calendar year. Existing subscribers are sent an invoice with the December Newsletter for the next year's subscription which offers a reduction if this is paid by the due date. If you are in arrears with your subscription a reminder notice comes attached to each issue of the Newsletter. Deadline for next issue The deadline for the December 1999 issue (Number 58) is 26 November 1999. Please forward contributions to: Dr Carol J. West, c/- Department of Conservation PO Box 743 Invercargill Contributions may be provided on an IBM compatible floppy disc (Word) or by e-mail to [email protected] Cover Illustration Plagiochila ramosissima with antheridial branches. -

South East 154 October 2019.Pdf
Australian Plants Society South East NSW Group Newsletter 154 October 2019 Corymbia maculata Spotted Gum and Contac ts: President, Dianne Clark, [email protected] Macrozamia communis Burrawang Secretary, Paul Hattersley [email protected] Newsletter editor, John Knight, [email protected] Group contact [email protected] Next Meeting Saturday 2nd November 2019 Plants and Vegetation of Montague Island At ERBG Staff Meeting Room 10am Paul Hattersley, APS Member and also a Volunteer Guide for Montague Island, had originally proposed to lead a tour onto the island to discuss relevant conservation issues. However, as insufficient members expressed interest in taking the boat trip, this had to be cancelled, and likewise therefore, the planned activities at Narooma. Plans for our next meeting have now changed and we will meet at ERBG staff meeting room at 10am for morning tea, followed by a presentation by Paul at 10.30am: "The plants and vegetation of Montague Island, seabird conservation, and the impacts of weeds and other human disturbance and activity". The history and heritage of Montague Island, including the light station, will also be weaved into the talk. Paul apologises to those who wanted to join the field trip on Montague Island, but with low numbers the economics of hiring the boat proved unviable. Following Paul’s presentation, we will conduct the Show and Tell session (please bring your garden plants in flower) and after lunch, open discussion, end of year notices, and a walk at gardens to see what has happened over the past 12 months. Members might consider dining at the Chefs Cap Café, recently opened in its new location by the lakeside. -
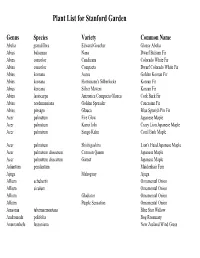
Plant List for Stanford Garden
Plant List for Stanford Garden Genus Species Variety Common Name Abelia grandiflora Edward Goucher Glossy Abelia Abies balsamea Nana Dwarf Balsam Fir Abies concolor Candicans Colorado White Fir Abies concolor Compacta Dwarf Colorado White Fir Abies koreana Aurea Golden Korean Fir Abies koreana Hortsmann's Silberlocke Korean Fir Abies koreana Silber Mavers Korean Fir Abies lasiocarpa Arizonica Compacta Glauca Cork Bark Fir Abies nordmanniana Golden Spreader Caucasian Fir Abies pinsapo Glauca Blue Spanish Pin Fir Acer palmatum Fire Glow Japanese Maple Acer palmatum Kurui Jishi Crazy Lion Japanese Maple Acer palmatum Sango Kaku Coral Bark Maple Acer palmatum Shishigashira Lion's Head Japanese Maple Acer palmatum dissectum Crimson Queen Japanese Maple Acer palmatum dissectum Garnet Japanese Maple Adiantum pendantum Maidenhair Fern Ajuga Mahogany Ajuga Allium schubertii Ornamental Onion Allium siculum Ornamental Onion Allium Gladiator Ornamental Onion Allium Purple Sensation Ornamental Onion Amsonia tabernaemontana Blue Star Willow Andromeda polifolia Bog Rosemary Anemanthele lessoniana New Zealand Wind Grass Genus Species Variety Common Name Anemone coronaria de Caen Wind Flower Anemone hupehensis japonica September Charm Japanese Anemone Anemone huphensis japonica Pamina Japanese Anemone Anemone x hybrida Honorine Jobert Japanese Anemone Angelica Aquilegia vulgaris Lime Frost Columbine Aquilegia Woodside Golden Columbine Aquillega Yellow Queen Yellow Columbine Arabis caucasica Variegata Variegated Rock Cress Arisaema formosanum DJHT 99049 Armeria maritima Alba White Sea Thrift Armeria maritima Dusseldorf Pride Sea Thrift Armeria pseudarmeria Arrhenatherum elatius ssp. Bulbosum Variegatum Striped Tuber Oat Grass Artemisia Oriental Limelinght Wormwood Artemisia Powis Castle Wormwood Arum italicum Wild Ginger Aruncus aethusifolius Dwarf Korean Goat's Beard Arundo donax Variegata Striped Giant Reed Arundo donax Giant Reed Asparagus densiflorus Myers Asparagus Fern/Pony Tail Fern Asplenium scolopendrium Hart's Tounge Fern Aster dumosus Prof A. -

Pollination Drop in Relation to Cone Morphology in Podocarpaceae: a Novel Reproductive Mechanism Author(S): P
Pollination Drop in Relation to Cone Morphology in Podocarpaceae: A Novel Reproductive Mechanism Author(s): P. B. Tomlinson, J. E. Braggins, J. A. Rattenbury Source: American Journal of Botany, Vol. 78, No. 9 (Sep., 1991), pp. 1289-1303 Published by: Botanical Society of America Stable URL: http://www.jstor.org/stable/2444932 . Accessed: 23/08/2011 15:47 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to American Journal of Botany. http://www.jstor.org AmericanJournal of Botany 78(9): 1289-1303. 1991. POLLINATION DROP IN RELATION TO CONE MORPHOLOGY IN PODOCARPACEAE: A NOVEL REPRODUCTIVE MECHANISM' P. B. TOMLINSON,2'4 J. E. BRAGGINS,3 AND J. A. RATTENBURY3 2HarvardForest, Petersham, Massachusetts 01366; and 3Departmentof Botany, University of Auckland, Auckland, New Zealand Observationof ovulatecones at thetime of pollinationin the southernconiferous family Podocarpaceaedemonstrates a distinctivemethod of pollencapture, involving an extended pollinationdrop. Ovules in all generaof the family are orthotropousand singlewithin the axil of each fertilebract. In Microstrobusand Phyllocladusovules are-erect (i.e., the micropyle directedaway from the cone axis) and are notassociated with an ovule-supportingstructure (epimatium).Pollen in thesetwo genera must land directly on thepollination drop in theway usualfor gymnosperms, as observed in Phyllocladus.In all othergenera, the ovule is inverted (i.e., the micropyleis directedtoward the cone axis) and supportedby a specializedovule- supportingstructure (epimatium). -

Newsletter #84 Winter 2015
NEWSLETTER WINTER 2015 FRIENDS OF THE WAITE ARBORETUM INC. NUMBER 84 www.waite.adelaide.edu.au/waite-historic/arboretum Patron: Sophie Thomson President: Beth Johnstone OAM, Vice-President: Marilyn Gilbertson OAM FORTHCOMING EVENTS Secretary: vacant, Treasurer: Dr Peter Nicholls FRIENDS OF THE WAITE Editor: Eileen Harvey, email: [email protected] ARBORETUM EVENTS Committee: Henry Krichauff, Robert Boardman, Norma Lee, Ron Allen, Dr Wayne Harvey, Terry Langham, Dr Jennifer Gardner (ex officio) Free Guided Arboretum walks Address: Friends of the Waite Arboretum, University of Adelaide, Waite Campus, The first Sunday of every month PMB1, GLEN OSMOND 5064 at 11.00 am. Phone: (08) 8313 7405, Email: [email protected] Walks meet at Urrbrae House Photography: Eileen Harvey Jacob and Gideon Cordover Guitar Concert, Wed. August 26 at Urrbrae House Ballroom. Refreshments 6 pm. 6.30 - 7.30 pm Performance of classical guitar and narration of the story of a little donkey and the simple joy of living. Enquiries and bookings please contact Beth Johnstone on 8357 1679 or [email protected] Spring visit to the historic house and grounds of Anlaby. Booking deadline extended. Unveiling of Bee Hotel Signage 11 am Tuesday 18 August Hakea francisiana, Grass-leaf Hakea More details at: http://www.adelaide.edu.au/ Table of contents waite-historic/whatson/ 2. From the President, Beth Johnstone OAM 3. From the Curator, Dr Jennifer Gardner. 4. Friends News: 5. Bee Hotel, Terry Langham. 6. Geijera parviflora, Wilga, Ron Allen. 7. Cork, Jean Bird. 8. Cork and the discovery of cells, Diarshul Sandhu. 9. The Conifers, Robert Boardman. -

Supporting Information
Supporting Information Mao et al. 10.1073/pnas.1114319109 SI Text BEAST Analyses. In addition to a BEAST analysis that used uniform Selection of Fossil Taxa and Their Phylogenetic Positions. The in- prior distributions for all calibrations (run 1; 144-taxon dataset, tegration of fossil calibrations is the most critical step in molecular calibrations as in Table S4), we performed eight additional dating (1, 2). We only used the fossil taxa with ovulate cones that analyses to explore factors affecting estimates of divergence could be assigned unambiguously to the extant groups (Table S4). time (Fig. S3). The exact phylogenetic position of fossils used to calibrate the First, to test the effect of calibration point P, which is close to molecular clocks was determined using the total-evidence analy- the root node and is the only functional hard maximum constraint ses (following refs. 3−5). Cordaixylon iowensis was not included in in BEAST runs using uniform priors, we carried out three runs the analyses because its assignment to the crown Acrogymno- with calibrations A through O (Table S4), and calibration P set to spermae already is supported by previous cladistic analyses (also [306.2, 351.7] (run 2), [306.2, 336.5] (run 3), and [306.2, 321.4] using the total-evidence approach) (6). Two data matrices were (run 4). The age estimates obtained in runs 2, 3, and 4 largely compiled. Matrix A comprised Ginkgo biloba, 12 living repre- overlapped with those from run 1 (Fig. S3). Second, we carried out two runs with different subsets of sentatives from each conifer family, and three fossils taxa related fi to Pinaceae and Araucariaceae (16 taxa in total; Fig. -

Podocarpus Has a fleshy Enlarged Stem with the Seed Capsule Attached
Some members of the Podocarpaceae It now seems as if the general lock down is being eased across the country and I am looking to do other things than write these notes which seemed helpful eleven weeks ago but hopefully soon will not be so. Not quite a bakers dozen but one more than a S.I. preferred unit. Also I must get on with the June FACTT Newsletter which I like to think is as eagerly awaited as these notes. This note is about a family, members of which are found in fossils of Gondwana times and now in those countries which formed the original landmass and some other countries. Podocarpus has a fleshy enlarged stem with the seed capsule attached. It is easy to see the possibility of birds spreading the seeds, unlike Nothofagus where the seed is less likely to be spread widely one would think. Regarding the movement of continents it needs to be mentioned that a million is a very large number. If, as Australia is presently doing, a landmass moves 70mm a year in just 1 million years; a short period of time in geological terms; the distance traveled is 70km. Australia is moving so fast in fact that GPS can’t keep up. Some people think that taxonomists fall into two broad categories - Lumpers and Splinters. Lumpers tend to say -Well the species are so similar they are in fact one. Splitters on the other hand tend to the view that the fine details represent sufficient differences to recognise they are different species. -

CUPRESSACEAE Actinostrobus Pyramidalis Swamp Cypress
CUPRESSACEAE Actinostrobus pyramidalis Swamp cypress Female cones Avon Catchment Council Actinostrobus pyramidalis Swamp cypress Plant features Growth form Erect, dense, narrowly pyramidal shaped shrub 1-4m tall. Leaves The leaves are bright dark green, triangular in cross section, with three distinctive longitudinal ridges, 0.5-2mm long and held CUPRESSACEAE either erect or slightly spreading close to the stem. Cones The male cone are cylindrical to egg shaped, appear on the end of branchlets and are 2-8mm x 1.5-2.5mm. The female cones are egg shaped to almost circular 10-15mm long, green with usually 6 valves running from the stem to the tip which are also egg shaped. On maturing the valves become brown with a ‘scale’ at the base of each valve that has a grey margin around a light brown band which has a dark brown area inside. The seeds are 5-4mm long, fawn in colour with three lighter coloured wings. The young female and male cones are present from Aug-Nov with the female cones lasting all year. Bark Coarse, fissured dark grey brown bark over the entire tree. Distribution Found from near Kalbarri to Ravensthorpe along the coastal plain Floodfringe and throughout the western half of Floodway the Avon catchment. Normal winter level Zone, habitat Prefered habitat of Actinostrobus pyramidalis Occurs in floodways of swamps, creeks, rivers, estuaries and other brackish to saline winter wet depressions. Grows on a sandy soil types that may contain clay or loam. Additional information An unusual shrub that is useful on a range of brackish to saline waterbodies. -

Gymnosperms on the EDGE Félix Forest1, Justin Moat 1,2, Elisabeth Baloch1, Neil A
www.nature.com/scientificreports OPEN Gymnosperms on the EDGE Félix Forest1, Justin Moat 1,2, Elisabeth Baloch1, Neil A. Brummitt3, Steve P. Bachman 1,2, Stef Ickert-Bond 4, Peter M. Hollingsworth5, Aaron Liston6, Damon P. Little7, Sarah Mathews8,9, Hardeep Rai10, Catarina Rydin11, Dennis W. Stevenson7, Philip Thomas5 & Sven Buerki3,12 Driven by limited resources and a sense of urgency, the prioritization of species for conservation has Received: 12 May 2017 been a persistent concern in conservation science. Gymnosperms (comprising ginkgo, conifers, cycads, and gnetophytes) are one of the most threatened groups of living organisms, with 40% of the species Accepted: 28 March 2018 at high risk of extinction, about twice as many as the most recent estimates for all plants (i.e. 21.4%). Published: xx xx xxxx This high proportion of species facing extinction highlights the urgent action required to secure their future through an objective prioritization approach. The Evolutionary Distinct and Globally Endangered (EDGE) method rapidly ranks species based on their evolutionary distinctiveness and the extinction risks they face. EDGE is applied to gymnosperms using a phylogenetic tree comprising DNA sequence data for 85% of gymnosperm species (923 out of 1090 species), to which the 167 missing species were added, and IUCN Red List assessments available for 92% of species. The efect of diferent extinction probability transformations and the handling of IUCN data defcient species on the resulting rankings is investigated. Although top entries in our ranking comprise species that were expected to score well (e.g. Wollemia nobilis, Ginkgo biloba), many were unexpected (e.g.