Citric Acid Cycle - O C O
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Effect of Citric Acid Cycle Genetic Variants and Their Interactions With
cancers Article Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank Sooyoung Cho 1 , Nan Song 2,3 , Ji-Yeob Choi 2,4,5 and Aesun Shin 1,2,* 1 Department of Preventive Medicine, Seoul National University College of Medicine, Seoul 03080, Korea; [email protected] 2 Cancer Research Institute, Seoul National University, Seoul 03080, Korea; [email protected] (N.S.); [email protected] (J.-Y.C.) 3 Department of Epidemiology and Cancer Control, St. Jude Children’s Research Hospital, Memphis, TN 38105, USA 4 Department of Biomedical Sciences, Graduate School of Seoul National University, Seoul 03080, Korea 5 Medical Research Center, Institute of Health Policy and Management, Seoul National University, Seoul 03080, Korea * Correspondence: [email protected]; Tel.: +82-2-740-8331; Fax: +82-2-747-4830 Received: 18 August 2020; Accepted: 9 October 2020; Published: 12 October 2020 Simple Summary: The citric acid cycle has a central role in the cellular energy metabolism and biosynthesis of macromolecules in the mitochondrial matrix. We identified the single nucleotide polymorphisms (SNPs) of the citrate acid cycle with colorectal cancer susceptibility in UK population. Furthermore, we found the significant interaction of SNPs in the citric acid cycle with the contributors to energy balance and SNP-SNP interactions. Our findings provide clues to the etiology in cancer development related to energy metabolism and evidence on identification of the population at high risk of colorectal cancer. -

Nutrition and Metabolism
NUTRITION AND METABOLISM Metabolism - the sum of the chemical changes that occur in the cell and involve the breakdown (catabolism) and synthesis (anabolism) of stored energy sources. Basal Metabolic Rate is dened as the rate of energy production by the body measured under a dened set of conditions which is usually at rest (physical and mental), room temperature, 12 hours after a meal. The result is produced as a percentage of a standard value which is derived from studies of normal healthy people. Measurement of the metabolic rate takes place using a method called calorimetry. This may be done directly by measuring the amount of heat produced by the body in an Atwater chamber, the metabolic rate is the amount of heat produced per hour. More commonly the metabolic rate is determined indirectly by putting people on a closed circuit breathing system, with CO2 removed by a soda lime scrubber and the rate of oxygen consumption measured by change in volume. Oxygen consumption is proportional to the metabolic rate because most of the energy in the body is derived from oxidative phosphorylation, which uses a set amount of oxygen to produce a set amount of energy. For every litre of oxygen consumed the body produces (uses) 4.82 kcals of energy. If the oxygen consumption is 250ml/min (15L/hr) then the metabolic rate is 72.3 kcals/hr. This is often further rened by dividing the gure by the body surface area which for a 70kg male is 1.73m2. This gives an average BMR of approximately 40 kcal/m2/hr. -

Tricarboxylic Acid (TCA) Cycle Intermediates: Regulators of Immune Responses
life Review Tricarboxylic Acid (TCA) Cycle Intermediates: Regulators of Immune Responses Inseok Choi , Hyewon Son and Jea-Hyun Baek * School of Life Science, Handong Global University, Pohang, Gyeongbuk 37554, Korea; [email protected] (I.C.); [email protected] (H.S.) * Correspondence: [email protected]; Tel.: +82-54-260-1347 Abstract: The tricarboxylic acid cycle (TCA) is a series of chemical reactions used in aerobic organisms to generate energy via the oxidation of acetylcoenzyme A (CoA) derived from carbohydrates, fatty acids and proteins. In the eukaryotic system, the TCA cycle occurs completely in mitochondria, while the intermediates of the TCA cycle are retained inside mitochondria due to their polarity and hydrophilicity. Under cell stress conditions, mitochondria can become disrupted and release their contents, which act as danger signals in the cytosol. Of note, the TCA cycle intermediates may also leak from dysfunctioning mitochondria and regulate cellular processes. Increasing evidence shows that the metabolites of the TCA cycle are substantially involved in the regulation of immune responses. In this review, we aimed to provide a comprehensive systematic overview of the molecular mechanisms of each TCA cycle intermediate that may play key roles in regulating cellular immunity in cell stress and discuss its implication for immune activation and suppression. Keywords: Krebs cycle; tricarboxylic acid cycle; cellular immunity; immunometabolism 1. Introduction The tricarboxylic acid cycle (TCA, also known as the Krebs cycle or the citric acid Citation: Choi, I.; Son, H.; Baek, J.-H. Tricarboxylic Acid (TCA) Cycle cycle) is a series of chemical reactions used in aerobic organisms (pro- and eukaryotes) to Intermediates: Regulators of Immune generate energy via the oxidation of acetyl-coenzyme A (CoA) derived from carbohydrates, Responses. -

Citric Acid Cycle
CHEM464 / Medh, J.D. The Citric Acid Cycle Citric Acid Cycle: Central Role in Catabolism • Stage II of catabolism involves the conversion of carbohydrates, fats and aminoacids into acetylCoA • In aerobic organisms, citric acid cycle makes up the final stage of catabolism when acetyl CoA is completely oxidized to CO2. • Also called Krebs cycle or tricarboxylic acid (TCA) cycle. • It is a central integrative pathway that harvests chemical energy from biological fuel in the form of electrons in NADH and FADH2 (oxidation is loss of electrons). • NADH and FADH2 transfer electrons via the electron transport chain to final electron acceptor, O2, to form H2O. Entry of Pyruvate into the TCA cycle • Pyruvate is formed in the cytosol as a product of glycolysis • For entry into the TCA cycle, it has to be converted to Acetyl CoA. • Oxidation of pyruvate to acetyl CoA is catalyzed by the pyruvate dehydrogenase complex in the mitochondria • Mitochondria consist of inner and outer membranes and the matrix • Enzymes of the PDH complex and the TCA cycle (except succinate dehydrogenase) are in the matrix • Pyruvate translocase is an antiporter present in the inner mitochondrial membrane that allows entry of a molecule of pyruvate in exchange for a hydroxide ion. 1 CHEM464 / Medh, J.D. The Citric Acid Cycle The Pyruvate Dehydrogenase (PDH) complex • The PDH complex consists of 3 enzymes. They are: pyruvate dehydrogenase (E1), Dihydrolipoyl transacetylase (E2) and dihydrolipoyl dehydrogenase (E3). • It has 5 cofactors: CoASH, NAD+, lipoamide, TPP and FAD. CoASH and NAD+ participate stoichiometrically in the reaction, the other 3 cofactors have catalytic functions. -
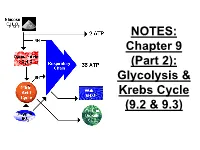
Chapter 9 (Part 2): Glycolysis & Krebs Cycle (9.2 & 9.3)
NOTES: Chapter 9 (Part 2): Glycolysis & Krebs Cycle (9.2 & 9.3) ● CELLULAR RESPIRATION: reactions in living cells in which sugars are broken down and energy is released Mitochondria in a Liver Cell!! Glucose + oxygen carbon dioxide + water + ENERGY C6H12O6 + 6O2 6CO2 + 6H2O + energy ● Food (glucose), like fuel, is “burned” by our cells for energy; however if it is burned all at once, too much energy is released. ● So, the reaction is broken down into many small steps controlled by ENZYMES ● the energy is transferred to the bonds of ATP which stores and releases the energy in usable amounts (packets) to be used by the cell Recall: the ATP cycle -Glucose = “large denomination” ($100) -ATP = “small change” ($1) *For each molecule of glucose, the cell can make approximately 36-38 ATP. Steps of Cellular Respiration: Phase of Occurs Starts Ends # of ATP Resp. where? with? with? made cyto- Glycoly- plasm 1 2 2 sis glucose pyruvate; NADH Krebs cycle (Citric Acid Cyc) E.T.C. (Resp Chain) & oxidative phosphor Steps of Cellular Respiration: Phase of Occurs Starts Ends # of ATP Resp. where? with? with? made cyto- Glycoly- plasm 1 2 2 sis glucose pyruvate; NADH Krebs inner 2 4 CO2, cycle matrix of pyruvate NADH, 2 (Citric Acid mito- FADH2 Cyc) chondria E.T.C. (Resp Chain) & oxidative phosphor Steps of Cellular Respiration: Phase of Occurs Starts Ends # of ATP Resp. where? with? with? made cyto- Glycoly- plasm 1 2 2 sis glucose pyruvate; NADH Krebs inner 2 4 CO2, cycle matrix of pyruvate NADH, 2 (Citric Acid mito- FADH2 Cyc) chondria E.T.C. -

Beta (Β)-Oxidation of Fatty Acid and Its Associated Disorders
Vol. 5 (1), pp. 158-172, December, 2018 ©Global Science Research Journals International Journal of Clinical Biochemistry Author(s) retain the copyright of this article. http://www.globalscienceresearchjournals.org/ Review Article Beta (β)-Oxidation of Fatty Acid and its associated Disorders Satyam Prakash Assistant Professor, Dept. of Biochemistry, Janaki Medical College Teaching Hospital, Janakpur, Nepal Mobile: +977-9841603704, E-mail: [email protected] Accepted 18 December, 2018 The lipids of metabolic significance in the mammalian organisms include triacylglycerols, phospholipids and steroids, together with products of their metabolism such as long-chain fatty acids, glycerol and ketone bodies. The fatty acids which are present in the triacylglycerols in the reduced form are the most abundant source of energy and provide energy twice as much as carbohydrates and proteins. Fatty acids represent an important source of energy in periods of catabolic stress related to increased muscular activity, fasting or febrile illness, where as much as 80% of the energy for the heart, skeletal muscles and liver could be derived from them. The prime pathway for the degradation of fatty acids is mitochondrial fatty acid β-oxidation (FAO). The relationship of fat oxidation with the utilization of carbohydrate as a source of energy is complex and depends upon tissue, nutritional state, exercise, development and a variety of other influences such as infection and other pathological states. Inherited defects for most of the FAO enzymes have been identified and characterized in early infancy as acute life-threatening episodes of hypoketotic, hypoglycemic coma induced by fasting or febrile illness. Therefore, this review briefly highlights mitochondrial β-oxidation of fatty acids and associated disorders with clinical manifestations. -

Glycolysis Citric Acid Cycle Oxidative Phosphorylation Calvin Cycle Light
Stage 3: RuBP regeneration Glycolysis Ribulose 5- Light-Dependent Reaction (Cytosol) phosphate 3 ATP + C6H12O6 + 2 NAD + 2 ADP + 2 Pi 3 ADP + 3 Pi + + 1 GA3P 6 NADP + H Pi NADPH + ADP + Pi ATP 2 C3H4O3 + 2 NADH + 2 H + 2 ATP + 2 H2O 3 CO2 Stage 1: ATP investment ½ glucose + + Glucose 2 H2O 4H + O2 2H Ferredoxin ATP Glyceraldehyde 3- Ribulose 1,5- Light Light Fx iron-sulfur Sakai-Kawada, F Hexokinase phosphate bisphosphate - 4e + center 2016 ADP Calvin Cycle 2H Stroma Mn-Ca cluster + 6 NADP + Light-Independent Reaction Phylloquinone Glucose 6-phosphate + 6 H + 6 Pi Thylakoid Tyr (Stroma) z Fe-S Cyt f Stage 1: carbon membrane Phosphoglucose 6 NADPH P680 P680* PQH fixation 2 Plastocyanin P700 P700* D-(+)-Glucose isomerase Cyt b6 1,3- Pheophytin PQA PQB Fructose 6-phosphate Bisphosphoglycerate ATP Lumen Phosphofructokinase-1 3-Phosphoglycerate ADP Photosystem II P680 2H+ Photosystem I P700 Stage 2: 3-PGA Photosynthesis Fructose 1,6-bisphosphate reduction 2H+ 6 ADP 6 ATP 6 CO2 + 6 H2O C6H12O6 + 6 O2 H+ + 6 Pi Cytochrome b6f Aldolase Plastoquinol-plastocyanin ATP synthase NADH reductase Triose phosphate + + + CO2 + H NAD + CoA-SH isomerase α-Ketoglutarate + Stage 2: 6-carbonTwo 3- NAD+ NADH + H + CO2 Glyceraldehyde 3-phosphate Dihydroxyacetone phosphate carbons Isocitrate α-Ketoglutarate dehydogenase dehydrogenase Glyceraldehyde + Pi + NAD Isocitrate complex 3-phosphate Succinyl CoA Oxidative Phosphorylation dehydrogenase NADH + H+ Electron Transport Chain GDP + Pi 1,3-Bisphosphoglycerate H+ Succinyl CoA GTP + CoA-SH Aconitase synthetase -

Cellular Respiration Liberation of Energy by Oxidation of Food
Cellular Respiration Liberation of Energy by Oxidation of Food Respiration and Photosynthesis: Photosynthesis uses CO2 and H2O molecules to form C6H12O6 (glucose) and O2. Respiration is just the opposite of photosynthesis; it uses O2 to breakdown glucose into CO2 and H2O. It results in chemical cycling in biosphere. Respiration and Breathing: Respiration takes place in cells and needs O2 to breakdown food and releases the waste matter CO2. Breathing exchanges these gases between lungs and air. Overall equation for cellular respiration is: C6H1206 + O2 6CO2 + 6 H2O + ATP Glucose Oxygen Carbon Dioxide Water Energy Redox reactions: reduction-oxidation reactions. The gain of electrons during a chemical reaction is called Reduction. The loss of electrons during a chemical reaction is called Oxidation. Glucose is oxidized to 6CO2 and O2 is reduced to 6H2O during cellular respiration. During cellular respiration, glucose loses electrons and H, and O2 gains them. Energy and Food All living things need energy. Some living things can make their food from CO2 and H2O – Producers (plants, algae) Animals feeding on plants – herbivores (chipmunk) Animals feeding on animals – Carnivores (lion) Producers change solar energy to chemical energy of organic molecules – glucose, amino acids Animals and also plants break chemical bonds of sugar molecules and make ATP. Use ATP for all cellular functions 4 Main Step of Cellular Respiration Glycolysis: Glucose + 2NAD + 2ADP 2 Pyruvate + 2NADH + 2 ATP Preparatory Step: Pyruvate + NAD Acetyl-CoA + CO2 + NADH Krebs Cycle: Acetyl-CoA + NAD + FAD + ADP CO2 + NADH + FADH + ATP Electron Transport Chain: electrons of NADH + O2 ATP + H2O Cellular Respiration Aerobic Harvest of energy: is the main source of energy for most organisms. -

Role of Hepatic Carbonic Anhydrase in De Novo Lipogenesis Christopher J
Biochem. J. (1995) 310,197-202 (Printed in Great Britain) 197 Role of hepatic carbonic anhydrase in de novo lipogenesis Christopher J. LYNCH,*t Heather FOX,* Stacy A. HAZEN,* Bruce A. STANLEY,* Susanna DODGSONt and Kathryn F. LANOUE* *Department of Cellular and Molecular Physiology, College of Medicine, The Milton S. Hershey Medical Center and Children's Hospital, The Pennsylvania State University, Hershey, PA 17033, U.S.A. and tDeparment of Physiology, University of Pennsylvania, School of Medicine, Philadelphia, PA 19104, U.S.A. The role of carbonic anhydrase in de novo lipid synthesis was incorporation of 14C into acetoacetate. Since ATP citrate-lyase, examined by measuring [1-14C]acetate incorporation into total the cytosolic enzyme that catalyses the conversion of citrate into lipids, fatty acids and non-saponifiable lipids in freshly isolated acetyl-CoA, catalyses an early rate-limiting step in fatty acid rat hepatocytes. Two carbonic anhydrase inhibitors, trifluoro- synthesis, levels ofcytosolic citrate may be rate controlling for de methylsulphonamide (TFMS) and ethoxozolamide (ETZ) novo fatty acid and sterol synthesis. Indeed citrate concentrations decreased incorporation of 14C into total lipids. Both fatty acid were significantly reduced to 37 + 6 % of control in hepatocytes and non-saponifiable lipid components of the total lipid were incubated with 100 #M TFMS for 30 min. TFMS also inhibited inhibited to approximately the same extent by 100 ,M TFMS the incorporation of 14C from [1-14C]pyruvate into malate, citrate (29 + 0.3 % and 35 + 0.3 % of control respectively in replicate and glutamate, but not into lactate. This supports the hypothesis studies). However, neither drug significantly affected ATP con- that TFMS inhibits pyruvate carboxylation, i.e. -

Lecture 9: Citric Acid Cycle/Fatty Acid Catabolism
Metabolism Lecture 9 — CITRIC ACID CYCLE/FATTY ACID CATABOLISM — Restricted for students enrolled in MCB102, UC Berkeley, Spring 2008 ONLY Bryan Krantz: University of California, Berkeley MCB 102, Spring 2008, Metabolism Lecture 9 Reading: Ch. 16 & 17 of Principles of Biochemistry, “The Citric Acid Cycle” & “Fatty Acid Catabolism.” Symmetric Citrate. The left and right half are the same, having mirror image acetyl groups (-CH2COOH). Radio-label Experiment. The Krebs Cycle was tested by 14C radio- labeling experiments. In 1941, 14C-Acetyl-CoA was used with normal oxaloacetate, labeling only the right side of drawing. But none of the label was released as CO2. Always the left carboxyl group is instead released as CO2, i.e., that from oxaloacetate. This was interpreted as proof that citrate is not in the 14 cycle at all the labels would have been scrambled, and half of the CO2 would have been C. Prochiral Citrate. In a two-minute thought experiment, Alexander Ogston in 1948 (Nature, 162: 963) argued that citrate has the potential to be treated as chiral. In chemistry, prochiral molecules can be converted from achiral to chiral in a single step. The trick is an asymmetric enzyme surface (i.e. aconitase) can act on citrate as through it were chiral. As a consequence the left and right acetyl groups are not treated equivalently. “On the contrary, it is possible that an asymmetric enzyme which attacks a symmetrical compound can distinguish between its identical groups.” Metabolism Lecture 9 — CITRIC ACID CYCLE/FATTY ACID CATABOLISM — Restricted for students enrolled in MCB102, UC Berkeley, Spring 2008 ONLY [STEP 4] α-Keto Glutarate Dehydrogenase. -

Insights Into the Hexose Liver Metabolism—Glucose Versus Fructose
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2017 Insights into the Hexose Liver Metabolism-Glucose versus Fructose Geidl-Flueck, Bettina ; Gerber, Philipp A Abstract: High-fructose intake in healthy men is associated with characteristics of metabolic syndrome. Extensive knowledge exists about the differences between hepatic fructose and glucose metabolism and fructose-specific mechanisms favoring the development of metabolic disturbances. Nevertheless, the causal relationship between fructose consumption and metabolic alterations is still debated. Multiple effects of fructose on hepatic metabolism are attributed to the fact that the liver represents the major sink of fructose. Fructose, as a lipogenic substrate and potent inducer of lipogenic enzyme expression, enhances fatty acid synthesis. Consequently, increased hepatic diacylglycerols (DAG) are thought to directly interfere with insulin signaling. However, independently of this effect, fructose may also counteract insulin-mediated effects on liver metabolism by a range of mechanisms. It may drive gluconeogenesis not only as a gluconeogenic substrate, but also as a potent inducer of carbohydrate responsive element binding protein (ChREBP), which induces the expression of lipogenic enzymes as well as gluconeogenic enzymes. It remains a challenge to determine the relative contributions of the impact of fructose on hepatic transcriptome, proteome and allosterome changes and consequently on the regulation of plasma glucose metabolism/homeostasis. Mathematical models exist modeling hepatic glucose metabolism. Fu- ture models should not only consider the hepatic adjustments of enzyme abundances and activities in response to changing plasma glucose and insulin/glucagon concentrations, but also to varying fructose concentrations for defining the role of fructose in the hepatic control of plasma glucose homeostasis. -

Regulation of Ketone Body and Coenzyme a Metabolism in Liver
REGULATION OF KETONE BODY AND COENZYME A METABOLISM IN LIVER by SHUANG DENG Submitted in partial fulfillment of the requirements For the Degree of Doctor of Philosophy Dissertation Adviser: Henri Brunengraber, M.D., Ph.D. Department of Nutrition CASE WESTERN RESERVE UNIVERSITY August, 2011 SCHOOL OF GRADUATE STUDIES We hereby approve the thesis/dissertation of __________________ Shuang Deng ____________ _ _ candidate for the ________________________________degree Doctor of Philosophy *. (signed) ________________________________________________ Edith Lerner, PhD (chair of the committee) ________________________________________________ Henri Brunengraber, MD, PhD ________________________________________________ Colleen Croniger, PhD ________________________________________________ Paul Ernsberger, PhD ________________________________________________ Janos Kerner, PhD ________________________________________________ Michelle Puchowicz, PhD (date) _______________________June 23, 2011 *We also certify that written approval has been obtained for any proprietary material contained therein. I dedicate this work to my parents, my son and my husband TABLE OF CONTENTS Table of Contents…………………………………………………………………. iv List of Tables………………………………………………………………………. viii List of Figures……………………………………………………………………… ix Acknowledgements………………………………………………………………. xii List of Abbreviations………………………………………………………………. xiv Abstract…………………………………………………………………………….. xvii CHAPTER 1: KETONE BODY METABOLISM 1.1 Overview……………………………………………………………………….. 1 1.1.1 General introduction