Beneficial Actions of Orostachys Japonica and Its Compounds Against Tumors Via MAPK Signaling Pathways
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Establishment of a Rapid and Efficient Micropropagation System For
HORTSCIENCE 52(9):1278–1282. 2017. doi: 10.21273/HORTSCI12056-17 been considered as an attractive way of propagating Haworthia genus plants. This method facilitates the propagation of plants Establishment of a Rapid and Efficient without seasonal and environment limitations and in a large scale, which are important Micropropagation System factors in the breeding plants for ornamental and landscaping purposes (Kitamura et al., for Succulent Plant Haworthia 2002; Kumari et al., 2016; Preil et al., 1988). Plantlets have been successfully induced in several Haworthia species using explants turgida Haw. from inflorescences (Kaul and Sabharwal, Boling Liu, Hongzhou Fang, Chaorong Meng, Ming Chen, 1972; Majumdar, 1970a; Ogihara, 1979; and Qingdong Chai Ogihara and Tsunewaki, 1978), ovary walls (Majumdar, 1970b), and leaves (Beyl and College of Life Sciences, Qufu Normal University, 273165, Qufu, Shandong, Sharma, 1983; Rogers, 1993b). Growth char- China acteristics in response to various auxins and cytokinins, such as NAA, 2,4-D, and BA Kai Zhang (Kaul and Sabharwal, 1972; Ogihara, 1979; Culaishan Forest Farm of Tai’an City, 271000, Tai’an, Shandong, China Ogihara and Tsunewaki, 1978), have been 1 examined in some Haworthia species. How- Shijuan Liu ever, none of these studies have used TDZ in College of Life Sciences, Qufu Normal University, 273165, Qufu, Shandong, in vitro propagation, although it is an efficient China PGR and is widely used for rapid propagation of plant tissues (Fatima and Anis, 2011; Additional index words. callus, plant growth regulators, propagation, root, shoot Kitamura et al., 2002; Kumari et al., 2016; Abstract. In the present study, the effect of plant growth regulators (PGRs) on callus Murthy et al., 1998). -

5. OROSTACHYS Fischer, Mém. Soc. Imp. Naturalistes Moscou 2: 270. 1809. 瓦松属 Wa Song Shu Herbs Biennial
Flora of China 8: 206–209. 2001. 5. OROSTACHYS Fischer, Mém. Soc. Imp. Naturalistes Moscou 2: 270. 1809. 瓦松属 wa song shu Herbs biennial. Roots fibrous. Rhizome absent. Leaves of 1st year arranged in a solitary, basal, dense rosette, alternate, linear to ovate, often with dull purple dots, apex usually cuspidate with a white, cartilaginous appendage to rarely softly obtuse or acuminate. Flowering stem solitary, arising from center of rosette in 2nd year; stem leaves alternate. Inflorescense terminal, a dense raceme or thyrse, narrowly pyramidal to cylindric in outline, with a distinct main axis and sometimes cymose lateral branches, many flowered, bracteate. Flowers bisexual, subsessile or pedicellate, 5-merous. Sepals usually shorter than petals. Petals subconnate at base, white, pink, or red, lanceolate. Stamens 2 × as many as petals, in 2 series. Nectar scales small. Carpels erect, free, stipitate, many ovuled. Styles slender. Follicles beaked at apex, many seeded. Thirteen species: China, Japan, Kazakstan, Korea, Mongolia, Russia; eight species (one endemic) in China. 1a. Apical spine of rosette leaves absent; bracts spatulate-ovate .................................................... 1. O. malacophylla 1b. Apical spine of rosette leaves cuspidate; bracts oblong to linear. 2a. Apical spine or appendage of rosette leaves not cartilaginous .................................................... 2. O. japonica 2b. Apical spine or appendage of rosette leaves cartilaginous. 3a. Rosette leaves apically fimbriate or spine 1-toothed on each side. 4a. Margin of rosette leaf appendage fimbriate; petals red or white ..................................... 3. O. fimbriata 4b. Margin of rosette leaf appendage entire, sometimes spine 1-toothed on each side; petals white ................................................................................................................................. 4. O. chanetii 3b. Rosette leaves apically fimbriate, sometimes ± undulate. -

(12) United States Plant Patent (10) Patent No.: US PP27,113 P2 Hansen (45) Date of Patent: Aug
USOOPP27113P2 (12) United States Plant Patent (10) Patent No.: US PP27,113 P2 Hansen (45) Date of Patent: Aug. 30, 2016 (54) XSEDORO PLANT NAMED 'BLUE ELF PUBLICATIONS (50) Latin Name: xSedoro USDA Plants Database Classification for Kingdom Plantae Downto Varietal Denomination: Blue Elf Genus Sedum L., http://plants.usda.gov/java/ ClassificationServlet?format print (71) Applicant: Christopher M. Hansen, Holland, MI &fileNamePF=classificationPF201602081.03235343.txt (US) &title=Sedum'9620L., pulled from the internet on Feb. 8, 2016.* (72) Inventor: Christopher M. Hansen, Holland, MI * cited by examiner (US) Primary Examiner — Anne Grunberg (*) Notice: Subject to any disclaimer, the term of this (74) Attorney, Agent, or Firm — Penny J. Aguirre patent is extended or adjusted under 35 U.S.C. 154(b) by 66 days. (57) ABSTRACT A new cultivar of xSedoro plant named Blue Elf character (21) Appl. No.: 14/121,759 ized by its compact, non-flopping, mound-forming, low growing groundcover plant habit, its compact and dense clus (22) Filed: Oct. 14, 2014 ters offlowers that cover almost the entire plant surface (95%) in late Summer, its numerous clusters of flowers that are hot (51) Int. Cl. pink in color in late Summer, its foliage that is powder-blue in AOIH 5/2 (2006.01) color during spring and early Summer and becoming lightly (52) U.S. Cl. suffused with rose-pink in mid-summer to fall, its heavy USPC ....................................................... Pt/263.1 self-branching without pinching from early spring through (58) Field of Classification Search mid-Summer, its numerous lateral branches that elongate USPC ................................................ Plt./479, 263.1 away from the central stem, and its inflorescences that have a See application file for complete search history. -

Downloaded from the National Center for (Takara) According to the Manufacturer’S Protocol
Appl Biol Chem (2018) 61(5):499–508 Online ISSN 2468-0842 https://doi.org/10.1007/s13765-018-0383-3 Print ISSN 2468-0834 ARTICLE Markers for distinguishing Orostachys species by SYBR Green-based real-time PCR and verification of their application in commercial O. japonica food products Jun An1 . Jun-Cheol Moon2 . Cheol Seong Jang1 Received: 23 April 2018 / Accepted: 12 July 2018 / Published online: 18 July 2018 Ó The Korean Society for Applied Biological Chemistry 2018 Abstract Human consumption of plant functional foods qPCR system for detecting Orostachys species in O. has been rapidly increasing owing to the health benefits japonica food products, O. japonica DNA was detected in they provide. In particular, in Korea, the plant Orostachys all eight commercial products tested, with low Ct values japonica has attracted attention for its anticancer and other (\ 20), whereas none of the other Orostachys species effects. Of the 12 established Orostachys species, only DNAs were detected, confirming that the tested foods three (viz., O. iwarenge, O. malacophyllus, and O. japon- contained only O. japonica. Therefore, developed primers ica) have been allowed for use as foods in Korea. In this and qPCR conditions would be useful for verifying the study, 12 species-specific primer sets based on single authenticity of commercial O. japonica food products. nucleotide polymorphisms of five chloroplast genes and one nuclear gene were developed to discriminate Oros- Keywords Commercial O. japonica foods Á Orostachys Á tachys species through quantitative real-time PCR (qPCR) Real-time PCR Á Species-specific DNA markers analysis with SYBR Green staining. -

Long Handout HARDY
GROWING WINTER-HARDY CACTI AND OTHER SUCCULENTS OUTDOORS IN WESTERN COLORADO Don I. Campbell - November 2004 (Revised April 2012) Xeriphytic plants are adapted to dry arid conditions. Some have adapted to cold climates and are able to survive freezing temperatures. Many are classified as succulents. Succulents are defined as any drought-resistant (xeriphyte) plant able to store water in its stem, leaves, or roots. Cacti, sedums, sempervivums, ice plants, and agaves are examples of plants that store water in their leaves, stems and roots. Sagebrush, rabbit brush, native grasses and yucca are examples of non-succulent drought resistant xeriphytic plants. They do not store water but have root systems and other survival strategies that allow them to survive on very limited water supplies. There is no universal agreement about which plants are considered succulent. The degree of fleshiness and where it occurs gives clues to a particular plants succulence and cultural requirements. Succulent plants occur throughout much of the world. Cacti, however, are native only to the Western Hemisphere. A distinguishing characteristic of cacti are the areoles which produce spines and flowers. All cacti are succulents but not all succulents are cacti. CULTURAL REQUIREMENTS Growing many cacti and other succulents outdoors in our area is relatively simple IF you select the right plants and IF you can satisfy the plants basic needs for soil, drainage, moisture, minerals, light, and protection from pests. DRAINAGE - The most important consideration is drainage. Do not allow water to collect around the base of plants. Cacti are more sensitive than most other hardy succulents. If, after a rain, you find standing water in your garden which does not either sink in or drain off quickly, you may want to raise the plants or rework the soil to improve drainage. -

A Molecular Phylogenetic Study of the Genus Phedimus for Tracing the Origin of “Tottori Fujita” Cultivars
plants Article A Molecular Phylogenetic Study of the Genus Phedimus for Tracing the Origin of “Tottori Fujita” Cultivars Sung Kyung Han 1 , Tae Hoon Kim 2 and Jung Sung Kim 1,* 1 Department of Forest Science, Chungbuk National University, Chungcheongbuk-do 28644, Korea; [email protected] 2 National Korea Forest Seed & Variety Center, Chungcheongbuk-do 27495, Korea; [email protected] * Correspondence: [email protected] Received: 23 January 2020; Accepted: 14 February 2020; Published: 17 February 2020 Abstract: It is very important to confirm and understand the genetic background of cultivated plants used in multiple applications. The genetic background is the history of crossing between maternal and paternal plants to generate a cultivated plant. If the plant in question was generated from a simple origin and not complicated crossing, we can easily confirm the history using a phylogenetic tree based on molecular data. This study was conducted to trace the origin of “Tottori Fujita 1gou” and “Tottori Fujita 2gou”, which are registered as cultivars originating from Phedimus kamtschaticus. To investigate the phylogenetic position of these cultivars, the backbone tree of the genus Phedimus needed to be further constructed because it retains inarticulate phylogenetic relationships among the wild species. We performed molecular phylogenetic analysis for P. kamtschaticus, Phedimus takesimensis, Phedimus aizoon, and Phedimus middendorffianus, which are assumed as the species of origin for “Tottori Fujita 1gou” and “Tottori Fujita 2gou”. The molecular phylogenetic tree based on the internal transcribed spacer (ITS) and psbA-trnH sequences showed the monophyly of the genus Phedimus, with P. takesimensis forming a single clade. However, P. -

Crassulaceae
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/227205999 Crassulaceae Chapter · April 2007 DOI: 10.1007/978-3-540-32219-1_12 CITATIONS READS 31 417 2 authors: Joachim Thiede Urs Eggli 88 PUBLICATIONS 183 CITATIONS 65 PUBLICATIONS 584 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Ecology and ecophysiology of desert plants in the Succulent Karoo, Namib, Negev, Sahara and other drylands View project Contributions to the succulent flora of Malawi View project All content following this page was uploaded by Joachim Thiede on 19 May 2017. The user has requested enhancement of the downloaded file. Crassulaceae 93 r- subfa- clade taxon distribution ::"spp.tribe mily family 5 Slnocrassu/a l EI t- to I Kungia l, , .r Meterostachys ä f f f;mnerate lsl to I F Orostachys Append. subs. I Hytotetephium ) t!_il'l Umbilicus Rhodiola I Pseudosedum I temoerate t Rhodiota atiu 1e Medit') i F] f ) l"l Phedimus I E_l Sempervivum Europe/N.East rytvum S. assyrlacum Near East [G] N S. mooneyifG] NE Africa l=l ; EItEI lo I Petrosedum Eurooe/Medit. I,l lll - l"l n- Aeonium S. ser. Pubescens [G] I t--l S. ser. Caerulea lGl INorthAfrica tl rl, ) S. ser. Monanthoidea [G] -{ ES Aichryson tsl .))t\ Monanthes Macaronesia l'l r- Aeonium ] E] 1e S. magel/ense[G] ! rP S. dasyphyllum [G] S. tydium l-t ic lGl l.l ae Rosularia Europe/ Mediterranean/ l'l S. sedoides l'l [G] 'Leuco- Near EasV tl S. -

Hortus Plant List 2021 for Website
PLANT COLLECTION INVENTORY PLANT. INFRASPE- COUNTRY Month/Ye GENUS HYBRID GENUS NAME SPECIES HYBRID SPECIES NAME CIFIC CULTIVAR NAME COMMON NAME ORIGIN HORTUS TAG # NOTES PROPERTY GARDENS 7/16 7/16 ACER GRISEUM PAPERBARK MAPLE UNITEDCHINA 16-54 ACER PENSYLVANICUM MOOSEWOOD STATES ON PROPERTY ACER RUBRUM ON PROPERTY AESCULUS BOTTLEBRUSH AESCULUS PARVIFLORA BOTTLEBRUSHBUCKEYE UNITED AESCULUS VAR. SEROTINA ROGERS BUCKEYE STATES MISLABELED AESCULUS PAVIA RED BUCKEYE 2012 AJUGA REPTANS BLACK SCALLOP BUGLEWEED 2012 AJUGA REPTANS PINK LIGHTENING THREE-LEAF 05/05 AKEBIA TRIFOLIATA PURPLE ROSE AKEBIA ASIAN 05/05 AKEBIA QUINATA DEEP PURPLE FIVE-LEAF AKEBIA ASIAN 5-14 2017 AKEBIA QUINATA SILVER BELLS FIVE-LEAF AKEBIA ASIAN ALLIUM AFLATUNENSE PURPLE SENSATION ALLIUM GIGANTEUM GLOBEMASTER ALLIUM MOUNT EVEREST 2009 ALLIUM MULTIBULBOSUM PORCELAIN BERRY AMPELOPSIS BREVIPEDUNCULATA ELEGANS VINE ARISAEMA DRACONITUM GREEN DRAGON AKA ARISTOLOCHIA DURIOR DUTCHMAN'S PIPE MACROPHYLLA ARUNCUS MISTY LACE AZALEA R.HINDOGIRI X R. YEDOENSE Jun-16 AZALEA MACRONULATUM CORNELL PINK 16-46 AZALEA NUDIFLORUM UNITED BETULA PAPYRIFERA HEARTLEAFPAPERBIRCH STATES ON PROPERTY 2012 BRUNNERA KING'S RANSOM SIBERIANBRUNNERA BRUNNERA MACROPHYLLA LOOKING GLASS BUGLOSS BRUNNERA MACROPHYLLA JACK FROST CALLICARPA DICHOTOMA ISSAI BEAUTYBERRY Sep-18 CALLICARPA DICHOTOMA VAR. ALBIFRUCTA 9/18 CALLICARPA X ‘PEARL GLAM’ 2009 CALYCANTHUS FLORIDUS ATHENS SWEETSHRUB 2014 CALYCANTHUS FLORIDUS PURPUREA SWEETSHRUB 2010 CALYCANTHUS FLORIDUS MICHAEL LINDSEY 2010 CALYCANTHUS CHINENSIS VENUS CALYCANTHUS -

Download Denver-Botanic-Gardens-Living
LIVING COLLECTIONS MANAGEMENT PLAN Updated and Approved: November 28, 2017 Previously Updated and Approved: August 2008 Living Collections Plan - Page 1 of 53 LIVING COLLECTIONS MANAGEMENT PLAN October, 2017 Purpose of the Plan The Living Collections at Denver Botanic Gardens support the Gardens’ mission of connecting people with plants, especially plants from the Rocky Mountain Region and similar regions around the world. Collections showcase the use of right plants in the right place, educating the public about horticulture in the semi-arid, steppe climate of the Rocky Mountain and Plains regions. The diverse collections ranging from alpine to tropical plants fulfill the mission through education and conservation messages, and by “providing delight and enlightenment to everyone”. Adhering to the standards and guidelines articulated in the Living Collections Management Policy, this Plan provides staff taking care of these collections coordinated and uniform direction to assist in the maintenance, expansion, refinement and development of the collections over the specified period of time. Collections Content The Living Collections are comprised of seven major collections: 1. Alpine 2. Amenity 3. Aquatic 4. Cactus and Succulents 5. Native 6. Steppe 7. Tropical In addition to these seven major collections, the living collections also comprises of a growing Bonsai collection (included under Amenity Collection) and two nationally accredited collections registered with the American Public Gardens Association Plant Collection Network – Quercus (Oak) collection and Alpines of the World collection. Guiding Principles The acquisition and care of the collections will be guided by the Gardens’ four core values: • Diversity • Relevance • Sustainability • Transformation Review and Revision of the Plan The Collections Management Plan will be reviewed as needed at a minimum of every five years to assess the implementation of the priorities and activities specified herein and determine measures to overcome challenges in achieving them. -
October 2020 · Page 1
GREEN DRAGON TALES, OCTOBER 2020 · PAGE 1 ADIRONDACK CHAPTER North American Rock Garden Society IN THIS ISSUE: October 17 Program Membership Letter from the Chair Upcoming ACNARGS Programs A Small Plot and a Long Story Calendar of Select Events & Programs Experiences with Ornamental Alliums About ACNARGS September’s Available Online About NARGS National Late Bloomers ACNARGS Board Members and Contacts Daphne Cuttings, Take Two October 2020 “Tours” to Colorado and British Columbia, October 17, 1:00 PM via Zoom Terry Humphries, Program Gardening on the Roof of the Rocky Mountains A Hillside Garden Nick Courtens, Nicola Ripley, Colin in Coastal British Columbia Lee, Melissa Ebone & Betty Ford David Sellars Alpine Gardens Staff GREEN DRAGON TALES, OCTOBER 2020 · PAGE 2 Look for an email with the Zoom meeting link to join this October 17, 1:00 PM program. The presentations we will enjoy were developed by NARGS in a series called ‘Taproot.’ We will convene via Zoom in an interactive meeting format. Information about joining us will also be posted to the Adirondack Chapter’s Facebook page. We will continue meeting through the fall via Zoom with hopes that stellar programming from far reaches Will make up for the limitations of social distancing. Our September meeting, co-sponsored with Cornell Botanic Gardens and was well-attended and ran without technical snags, as far as we know. (If you have any issues or questions regarding Zoom, please let me know at [email protected].) Thanks should go to Carol Eichler for tapping contacts she formed organizing the NARGS conference to arrange CBG’s hosting of that Zoom Webinar with two great presentations by CBG staff. -

Pollen Morphology of the Family Crassulaceae from Pakistan and Kashmir and Its Taxonomic Implications
Pak. J. Bot. 47(4): 1481-1493, 2015. POLLEN MORPHOLOGY OF THE FAMILY CRASSULACEAE FROM PAKISTAN AND KASHMIR AND ITS TAXONOMIC IMPLICATIONS MUHAMMAD QAISER, ANJUM PERVEEN AND GHULAM RASOOL SARWAR Centre for Plant Conservation, University of Karachi, Karachi-75270, Pakistan Abstract Pollen morphology of 28 species representing 9 genera viz., Bryophyllum, Kalanchoe, Pseudosedum, Rhodiola, Rosularia, Hylotelephium, Sedum, Tillaea and Orostachys of the family Crassulaceae from Pakistan has been studied by light and scanning electron microscope. Pollen grains are usually free, radially symmetrical, isopolar, sub-oblate to oblate- spheroidal, or prolate–spheroidal to subprolate often prolate and tricolporate. Tectum mostly striate-rugulate or sub-psilate rarely reticulate-rugulate. Significant variation is found in shape and exine ornamentation and on the basis these characters family has been divided into three pollen types’ viz., Rhodiola sinuata–type, Rosularia viguieri–type and Sedum hispanicum–type. Numerical analysis of based on pollen characters of 28 species was also carried out to determine the phylogenetic relationships of various genera and the species of the family Crassulaceae. Key words: Crassulaceae, Eurapalynous, Exine ornamentation and Pollen shape. Introduction intergenic region (cpDNA) in Sedum (Crassulaceae) and geographic distribution. There are no reports on the pollen Crassulaceae, a family of about 33 genera and c. 1500 morphology of the family Crassulaceae from Pakistan and species is distributed all over the world, more Kashmir. However, Numarical taxonomy of the Sedum and predominantly in the temperate and subtropical zones of Rosularia from Pakistan has been studied by Sarwar & the northern hemisphere and Africa (Mabberely, 1987). Qaiser (2011, 2012) respectively. Present study is the first Represented in Pakistan by 9 genera and 32 species study of pollen morphology of the family Crassulaceae from (Sarwar, 2002). -
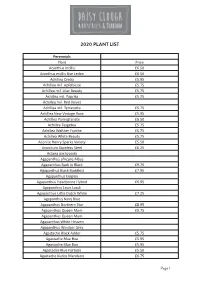
2020 Plant List
2020 PLANT LIST Perennials Plant Price Acanthus mollis £6.50 Acanthus mollis Rue Ledan £6.50 Achillea Credo £5.95 Achillea mil. Apfelbiute £5.75 Achillea mil. Lilac Beauty £5.75 Achillea mil. Paprika £5.75 Achillea mil. Red Velvet Achillea mil. Terracotta £5.75 Achillea New Vintage Rose £5.95 Achillea Pomegranate £6.50 Achillea Taygetea £5.75 Achillea Walther Funcke £5.75 Achillea White Beauty £5.75 Aconite Henry Sparks Variety £5.50 Aconitum Stainless Steel £6.25 Actaea pachypoda Agapanthus africans Albus Agapanthus Back in Black £9.75 Agapanthus Black Buddhist £7.95 Agapanthus Enigma Agapanthus Headborne Hybrid £6.95 Agapanthus Lapis Lazuli Agapanthus Little Dutch White £7.25 Agapanthus Navy Blue Agapanthus Northern Star £8.95 Agapanthus Queen Mum £9.75 Agapanthus Queen Mum Agapanthus White Heaven Agapanthus Windsor Grey Agastache Black Adder £5.75 Agastache Blue Boa £5.95 Agastache Blue Boa £5.95 Agastache Blue Fortune £5.50 Agastache Kudos Mandarin £6.75 Page 1 2020 PLANT LIST 2020 PLANT LIST Perennials Perennials Agastache Little Adder £6.75 Plant Price Agastache Red Fortune £5.95 Aruncus dioicus Kneiffii Alchemilla alpina Aruncus Guinea Fowl £7.25 Alchemilla erythropoda Asarum europaenum Alchemilla mollis Asclepius incarnata Alchemilla sericata Goldstrike Asclepius incarnata Soulmate £5.50 Allium Summer Beauty £5.95 Asphodeline lutea Alstroemeria Indian Summer £11.95 Asphodelus albus Alstroemeria Mona Lisa Aster King George £6.75 Amarine belladiva Anastasia Aster lateriflorus Lady in Black Amsonia tavern. Storm Cloud £6.50 Aster Little Carlow £5.95 Anaphalis triplinervis Aster maellus Veilchen Unigin £6.50 Anchusa azurea Dropmore £5.75 Aster Monch Anemone blanda White Splendour Aster novi -b.