2011 Management Indicator Species Assessment
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Notes on Italian Heptageniidae (Ephemeroptera). Rhithrogena Fiorii Grandi, 1953 and R
Aquatic Insects, Vol. 5 (1983), No. 2, pp. 69-76. Notes on Italian Heptageniidae (Ephemeroptera). Rhithrogena fiorii Grandi, 1953 and R. adrianae sp. n. by Carlo BELFIORE (Roma) ABSTRACT Rhithrogena adrianae, a new species related to R. diaphana Nav., is described from nymphs and male imagines collected in Central Italy. Taxonomic characters of nymphs and males of R. fiorii Grandi, whose nymphal stage was previously unknown, are also described and figured. Lectotype is designated for R. fiorii. The taxonomic status of Rhithrogena fiorii Grandi, 1953, described from winged stages only, was till now very uncertain. The type locality, near Bologna, is now altered by buildings and factories: R. fiorii has probably disappeared from that site. I have examined in Grandi's collection the specimens referred by her to R. fiorii, labelled: "Bologna, S. Luca, 16.III.1952 (l >, l < subim.), 20.III.1954 (l <, l > subim, l < subim.), 20.11.1955 (1 > subim.), 17.III.1955 (l <), .IV. 1955 (l >).I designate lectotype the male imago collected on 16.III. 1952. None of the spe- cimens is in a good state of preservation. Titillators are not truncate (Grandi, 1960: fig. 21,6 and pag. 91), but with few pointed lobes at the apex. During the first months of 1980 and 1981, in the river Mignone, near Rome, I collected and reared a hundred nymphs of Rhithrogena, from which I obtained some subimagines and two male imagines, easily referable to R. fiorii. I describe herein the taxonomic features of nymphs and males of this species. I also describe the male imago and nymph of a new species of Rhithrogena which lives in the same localities as R. -

CT DEEP Family-Level Identification Guide for Riffle-Dwelling Macroinvertebrates of Connecticut
CT DEEP Family-Level Identification Guide for Riffle-Dwelling Macroinvertebrates of Connecticut Seventh Edition Spring 2013 Authors and Acknowledgements Michael Beauchene produced the First Edition and revised the Second and Third Editions. Christopher Sullivan revised the Fourth and Fifth Editions. Erin McCollum developed the Sixth Edition with editorial assistance from Michael Beauchene. The First through Sixth Editions were developed and revised for use with Project SEARCH, a program formerly coordinated by CTDEEP but presently inactive. This Seventh Edition has been slightly modified for use by Connecticut high school students participating in the Connecticut Envirothon Aquatic Ecology workshop. Original drawings provided by Michael Beauchene and by the Volunteer Stream Monitoring Partnership at the University of Minnesota’s Water Resources Center. This page intentionally left blank. About the Key Scope of the Key This key is intended to assist Connecticut Envirothon students in the identification of aquatic benthic macroinvertebrates. As such, it is targeted toward organisms that are most commonly found in the riffle microhabitats of Connecticut streams. When conducting an actual field study of riffle dwelling macroinvertebrates, there may be an organism collected at a site in Connecticut that will not be found in this key. In this case, you should utilize another reference guide to identify the organism. Several useful guides are listed below. AQUATIC ENTOMOLOGY by Patrick McCafferty A GUIDE TO COMMON FRESHWATER INVERTEBRATES OF NORTH AMERICA by J. Reese Voshell, Jr. AN INTRODUCTION TO THE AQUATIC INSECTS OF NORTH AMERICA by R.W. Merritt and K.W. Cummins Most organisms will be keyed to the family level, however several will not be identified beyond the Kingdom Animalia phylum, class, or order. -

Lazare Botosaneanu ‘Naturalist’ 61 Doi: 10.3897/Subtbiol.10.4760
Subterranean Biology 10: 61-73, 2012 (2013) Lazare Botosaneanu ‘Naturalist’ 61 doi: 10.3897/subtbiol.10.4760 Lazare Botosaneanu ‘Naturalist’ 1927 – 2012 demic training shortly after the Second World War at the Faculty of Biology of the University of Bucharest, the same city where he was born and raised. At a young age he had already showed interest in Zoology. He wrote his first publication –about a new caddisfly species– at the age of 20. As Botosaneanu himself wanted to remark, the prominent Romanian zoologist and man of culture Constantin Motaş had great influence on him. A small portrait of Motaş was one of the few objects adorning his ascetic office in the Amsterdam Museum. Later on, the geneticist and evolutionary biologist Theodosius Dobzhansky and the evolutionary biologist Ernst Mayr greatly influenced his thinking. In 1956, he was appoint- ed as a senior researcher at the Institute of Speleology belonging to the Rumanian Academy of Sciences. Lazare Botosaneanu began his career as an entomologist, and in particular he studied Trichoptera. Until the end of his life he would remain studying this group of insects and most of his publications are dedicated to the Trichoptera and their environment. His colleague and friend Prof. Mar- cos Gonzalez, of University of Santiago de Compostella (Spain) recently described his contribution to Entomolo- gy in an obituary published in the Trichoptera newsletter2 Lazare Botosaneanu’s first contribution to the study of Subterranean Biology took place in 1954, when he co-authored with the Romanian carcinologist Adriana Damian-Georgescu a paper on animals discovered in the drinking water conduits of the city of Bucharest. -

(Trichoptera: Glossosomatidae: Protoptilinae) from Brazil
A new species of Protoptila Banks (Trichoptera: Glossosomatidae: Protoptilinae) from Brazil Allan Paulo Moreira SANTOS1, Jorge Luiz NESSIMIAN2 ABSTRACT A new species of Protoptila Banks (Trichoptera: Glossosomatidae: Protoptilinae) – P. longispinata sp. nov. – is described and illustrated from specimens collected in Amazon region, Amazonas and Pará states, Brazil. KEY WORDS: Amazon basin, Protoptila longispinata sp. nov., Neotropical Region, taxonomy. Uma nova espécie de Protoptila Banks (Trichoptera: Glossosomatidae: Protoptilinae) do Brasil RESUMO Uma nova espécie de Protoptila Banks (Trichoptera: Glossosomatidae: Protoptilinae) – P. longispinata sp. nov. – é descrita e ilustrada a partir de espécimes coletados na Região Amazônica, estados do Amazonas e do Pará, Brasil. PALAVRAS-CHAVE: bacia Amazônica, Protoptila longispinata sp. nov., Região Neotropical, taxonomia. 1 Universidade Federal do Rio de Janeiro. E-mail: [email protected] 2 Universidade Federal do Rio de Janeiro. E-mail: [email protected] 723 VOL. 39(3) 2009: 723 - 726 A new species of Protoptila Banks (Trichoptera: Glossosomatidae: Protoptilinae) from Brazil INTRODUCTION internal area slightly expanded. Forewings covered by long The genus Protoptila currently has 93 described species dark brown setae, and with a light transverse bar at midlength; widespread throughout the Americas, but with most species forks I, II, and III present; discoidal cell closed (Figure 1). occurring in the Neotropics (Robertson & Holzenthal, 2008). Hind wing with forks II and III present (Figure 2); nygma This is the largest genus of the subfamily Protoptilinae, and thyridium inconspicuous in fore- and hind wings. Legs represented in Brazil by 12 species, ten of which were described yellowish brown, with short dark setae. Abdominal segments from Amazon basin, nine occurring in Amazonas State: P. -

(Trichoptera: Limnephilidae) in Western North America By
AN ABSTRACT OF THE THESIS OF Robert W. Wisseman for the degree of Master ofScience in Entomology presented on August 6, 1987 Title: Biology and Distribution of the Dicosmoecinae (Trichoptera: Limnsphilidae) in Western North America Redacted for privacy Abstract approved: N. H. Anderson Literature and museum records have been reviewed to provide a summary on the distribution, habitat associations and biology of six western North American Dicosmoecinae genera and the single eastern North American genus, Ironoquia. Results of this survey are presented and discussed for Allocosmoecus,Amphicosmoecus and Ecclisomvia. Field studies were conducted in western Oregon on the life-histories of four species, Dicosmoecusatripes, D. failvipes, Onocosmoecus unicolor andEcclisocosmoecus scvlla. Although there are similarities between generain the general habitat requirements, the differences or variability is such that we cannot generalize to a "typical" dicosmoecine life-history strategy. A common thread for the subfamily is the association with cool, montane streams. However, within this stream category habitat associations range from semi-aquatic, through first-order specialists, to river inhabitants. In feeding habits most species are omnivorous, but they range from being primarilydetritivorous to algal grazers. The seasonal occurrence of the various life stages and voltinism patterns are also variable. Larvae show inter- and intraspecificsegregation in the utilization of food resources and microhabitatsin streams. Larval life-history patterns appear to be closely linked to seasonal regimes in stream discharge. A functional role for the various types of case architecture seen between and within species is examined. Manipulation of case architecture appears to enable efficient utilization of a changing seasonal pattern of microhabitats and food resources. -

Life History and Production of Mayflies, Stoneflies, and Caddisflies (Ephemeroptera, Plecoptera, and Trichoptera) in a Spring-Fe
Color profile: Generic CMYK printer profile Composite Default screen 1083 Life history and production of mayflies, stoneflies, and caddisflies (Ephemeroptera, Plecoptera, and Trichoptera) in a spring-fed stream in Prince Edward Island, Canada: evidence for population asynchrony in spring habitats? Michelle Dobrin and Donna J. Giberson Abstract: We examined the life history and production of the Ephemeroptera, Plecoptera, and Trichoptera (EPT) commu- nity along a 500-m stretch of a hydrologically stable cold springbrook in Prince Edward Island during 1997 and 1998. Six mayfly species (Ephemeroptera), 6 stonefly species (Plecoptera), and 11 caddisfly species (Trichoptera) were collected from benthic and emergence samples from five sites in Balsam Hollow Brook. Eleven species were abundant enough for life-history and production analysis: Baetis tricaudatus, Cinygmula subaequalis, Epeorus (Iron) fragilis,andEpeorus (Iron) pleuralis (Ephemeroptera), Paracapnia angulata, Sweltsa naica, Leuctra ferruginea, Amphinemura nigritta,and Nemoura trispinosa (Plecoptera), and Parapsyche apicalis and Rhyacophila brunnea (Trichoptera). Life-cycle timing of EPT taxa in Balsam Hollow Brook was generally similar to other literature reports, but several species showed extended emergence periods when compared with other studies, suggesting a reduction in synchronization of life-cycle timing, pos- sibly as a result of the thermal patterns in the stream. Total EPT secondary production (June 1997 to May 1998) was 2.74–2.80 g·m–2·year–1 dry mass (size-frequency method). Mayflies were dominant, with a production rate of 2.2 g·m–2·year–1 dry mass, followed by caddisflies at 0.41 g·m–2·year–1 dry mass, and stoneflies at 0.19 g·m–2·year–1 dry mass. -
The Study of the Zoobenthos of the Tsraudon River Basin (The Terek River Basin)
E3S Web of Conferences 169, 03006 (2020) https://doi.org/10.1051/e3sconf/202016903006 APEEM 2020 The study of the zoobenthos of the Tsraudon river basin (the Terek river basin) Ia E. Dzhioeva*, Susanna K. Cherchesova , Oleg A. Navatorov, and Sofia F. Lamarton North Ossetian state University named after K.L. Khetagurov, Vladikavkaz, Russia Abstract. The paper presents data on the species composition and distribution of zoobenthos in the Tsraudon river basin, obtained during the 2017-2019 research. In total, 4 classes of invertebrates (Gastropoda, Crustacea, Hydracarina, Insecta) are found in the benthic structure. The class Insecta has the greatest species diversity. All types of insects in our collections are represented by lithophilic, oligosaprobic fauna. Significant differences in the composition of the fauna of the Tsraudon river creeks and tributary streams have been identified. 7 families of the order Trichoptera are registered in streams, and 4 families in the river. It is established that the streamlets of the family Hydroptilidae do not occur in streams, the distribution boundary of the streamlets of Hydropsyche angustipennis (Hydropsychidae) is concentrated in the mountain-forest zone. The hydrological features of the studied watercourses are also revealed. 1 Introduction The biocenoses of flowing reservoirs of the North Caucasus, and especially small rivers, remain insufficiently explored today; particularly, there is no information about the systematic composition, biology and ecology of amphibiotic insects (mayflies, stoneflies, caddisflies and dipterous) of the studied basin. Amphibiotic insects are an essential link in the food chain of our reservoirs and at the same time can be attributed to reliable indicators of water quality. -

Resource Partitioning by Two Species of Stream Mayflies (Ephemeroptera: Heptageniidae)
The Great Lakes Entomologist Volume 14 Number 3 - Fall 1981 Number 3 - Fall 1981 Article 5 October 1981 Resource Partitioning by Two Species of Stream Mayflies (Ephemeroptera: Heptageniidae) William O. Lamp Illinois Natural History Survey N. Wilson Britt Ohio State University Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Lamp, William O. and Britt, N. Wilson 1981. "Resource Partitioning by Two Species of Stream Mayflies (Ephemeroptera: Heptageniidae)," The Great Lakes Entomologist, vol 14 (3) Available at: https://scholar.valpo.edu/tgle/vol14/iss3/5 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Lamp and Britt: Resource Partitioning by Two Species of Stream Mayflies (Ephemero 1981 THE GREAT LAKES ENTOMOLOGIST 151 RESOURCE PARTITIONING BY TWO SPECIES QF STREAM MAYFLIES (EPHEMEROPTERA: HEPTAGENIIDAE) William O. Lampl and N. Wilson Britt2 ABSTRACT We compared the phenology of nymph development, food type, and habitat selection of two stream mayflies, Stenacron interpunctatum (Say) and Stenonema pulchellum (Walsh) in Big Darby Creek, Ohio. Both species, which grow principally from autumn through early spring, emerged from the stream throughout the summer. The nymphs consumed the same sizes and types of food particles from deposits on stones, mostly in the form of detritus. As a result of morphological and behavioral adaptations, S. pulchellum lived on stones in swift water whereas S. -

A Comparison of Aquatic Invertebrate Assemblages Collected from the Green River in Dinosaur National Monument in 1962 and 2001
A Comparison of Aquatic Invertebrate Assemblages Collected from the Green River in Dinosaur National Monument in 1962 and 2001 Final Report for United States Department of the Interior National Park Service Dinosaur National Monument 4545 East Highway 40 Dinosaur, Colorado 81610-9724 Report Prepared by: Dr. Mark Vinson, Ph.D. & Ms. Erin Thompson National Aquatic Monitoring Center Department o f Fisheries and Wildlife Utah State University Logan, Utah 84322-5210 www.usu.edu/buglab 22 January 2002 i Foreword The work described in this report was conducted by personnel of the National Aquatic Monitoring Center, Utah State University, Logan, Utah. Mr. J. Matt Tagg aided in the identification of the aquatic invertebrates. Ms. Leslie Ogden provided computer assistance. Several people at Dinosaur National Monument helped us immensely with various project details. Steve Petersburg, Dana Dilsaver, and Dennis Ditmanson provided us with our research permit. Ann Elder helped with sample archiving. Christy Wright scheduled our trip and provided us with our river permit. We thank them all for all their help and good spirit. We also thank Mr. Walter Kittams (National Park Service, Regional Office, Omaha Nebraska) and Mr. Earl M. Semingsen (Superintendent, Dinosaur National Monument) for funding the study and the members of the 1962 University of Utah expedition: Dr. Angus M. Woodbury, Dr. Stephen Durrant, Mr. Delbert Argyle, Mr. Douglas Anderson, and Dr. Seville Flowers for their foresight to conduct the original study nearly 40 years ago. The concept and value of long-term ecological data is often bantered about, but its value is never more apparent then when we conduct studies like that presented here. -
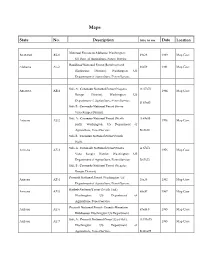
State No. Description Size in Cm Date Location
Maps State No. Description Size in cm Date Location National Forests in Alabama. Washington: ALABAMA AL-1 49x28 1989 Map Case US Dept. of Agriculture, Forest Service. Bankhead National Forest (Bankhead and Alabama AL-2 66x59 1981 Map Case Blackwater Districts). Washington: US Department of Agriculture, Forest Service. Side A : Coronado National Forest (Nogales A: 67x72 ARIZONA AZ-1 1984 Map Case Ranger District). Washington: US Department of Agriculture, Forest Service. B: 67x63 Side B : Coronado National Forest (Sierra Vista Ranger District). Side A : Coconino National Forest (North A:69x88 Arizona AZ-2 1976 Map Case Half). Washington: US Department of Agriculture, Forest Service. B:69x92 Side B : Coconino National Forest (South Half). Side A : Coronado National Forest (Sierra A:67x72 Arizona AZ-3 1976 Map Case Vista Ranger District. Washington: US Department of Agriculture, Forest Service. B:67x72 Side B : Coronado National Forest (Nogales Ranger District). Prescott National Forest. Washington: US Arizona AZ-4 28x28 1992 Map Case Department of Agriculture, Forest Service. Kaibab National Forest (North Unit). Arizona AZ-5 68x97 1967 Map Case Washington: US Department of Agriculture, Forest Service. Prescott National Forest- Granite Mountain Arizona AZ-6 67x48.5 1993 Map Case Wilderness. Washington: US Department of Agriculture, Forest Service. Side A : Prescott National Forest (East Half). A:111x75 Arizona AZ-7 1993 Map Case Washington: US Department of Agriculture, Forest Service. B:111x75 Side B : Prescott National Forest (West Half). Arizona AZ-8 Superstition Wilderness: Tonto National 55.5x78.5 1994 Map Case Forest. Washington: US Department of Agriculture, Forest Service. Arizona AZ-9 Kaibab National Forest, Gila and Salt River 80x96 1994 Map Case Meridian. -

Zootaxa, Canoptila (Trichoptera: Glossosomatidae)
CORE Metadata, citation and similar papers at core.ac.uk Provided by University of Minnesota Digital Conservancy Zootaxa 1272: 45–59 (2006) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA 1272 Copyright © 2006 Magnolia Press ISSN 1175-5334 (online edition) The Neotropical caddisfly genus Canoptila (Trichoptera: Glossosomatidae) DESIREE R. ROBERTSON1 & RALPH W. HOLZENTHAL2 University of Minnesota, Department of Entomology, 1980 Folwell Ave., Room 219, St. Paul, Minnesota 55108, U.S.A. E-mail: [email protected]; [email protected] ABSTRACT The caddisfly genus Canoptila Mosely (Glossosomatidae: Protoptilinae), endemic to southeastern Brazil, is diagnosed and discussed in the context of other protoptiline genera, and a brief summary of its taxonomic history is provided. A new species, Canoptila williami, is described and illustrated, including a female, the first known for the genus. Additionally, the type species, Canoptila bifida Mosely, is redescribed and illustrated. There are three possible synapomorphies supporting the monophyly of Canoptila: 1) the presence of long spine-like posterolateral processes on tergum X; 2) the highly membranous digitate parameres on the endotheca; and 3) the unique combination of both forewing and hind wing venational characters. Key words: Trichoptera, Glossosomatidae, Protoptilinae, Canoptila, new species, caddisfly, male genitalia, female genitalia, Neotropics, Atlantic Forest, southeastern Brazil INTRODUCTION The Atlantic Forest of southeastern Brazil is well known for its highly endemic flora and fauna, and has been designated a biodiversity hotspot (da Fonseca 1985; Myers et al. 2000). The forest, consisting of tropical evergreen and semideciduous mesophytic broadleaf species, originally covered most of the slopes of the coastal mountains and extended from well inland to the coastline (Fig. -

Final Report on Field Surveys
Ecological Surveys and Assessments to Facilitate Restoration Activities at the Salt River Marsh: Final Report on Field Surveys Prepared by: Peter J. Badra, Tyler J. Bassett, and Yu Man Lee Michigan Natural Features Inventory P.O. Box 13036 Lansing, MI 48901 For: Michigan Department of Environment, Great Lakes, and Energy, Water Resources Division February 4, 2020 Report Number 2020-05 Suggested Citation: Badra, P.J., T.J. Basset, Y.M. Lee. 2020. Ecological Surveys and Assessments to Facilitate Restoration Activities at the Salt River Marsh: Final Report on Field Surveys. Michigan Natural Features Inventory Report Number 2020-05, Lansing, MI. 97pp. Cover Photos: Background - Looking northwest at Stand 4, from edge of Stand 5, at cattails and common reed in the Salt River Marsh State Wildlife Area. Photo by Tyler J. Basset; Inset, left - Painted turtle hatchling. Photo by Yu Man Lee; Inset, right - American bluet damselfly larvae. Photo by Peter J. Badra. MSU is an affirmative-action, equal-opportunity employer. Michigan State University Extension programs and materials are open to all without regard to race, color, national origin, gender, gender identity, religion, age, height, weight, disability, political beliefs, sexual orientation, marital status, family status or veteran status. Copyright 2020 MSU Board of Trustees Acknowledgments Financial support for this project was provided by the Great Lakes Restoration Initiative via the Water Resources Division of the Michigan Department of Environment, Great Lakes, and Energy. Daria Hyde and Ashley Cole-Wick provided essential assistance with the field component of this project. Thank you to the landowners who kindly allowed us to access the Salt River and marsh through their properties, especially during difficult times with high water.