Anatomical Basis for Camouflaged Polarized Light Communication In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diffractive Properties of Blue Morpho Butterfly Wings
Diffractive Properties of Blue Morpho Butterfly Wings Mary Lalak and Paul Brackman Department of Physics and Astronomy, University of Georgia, Athens, Georgia 30602 (Dated: October 24, 2014) Many species of butterflies are known to produce beautiful iridescent colors when exposed to light from different angles. These affects can be attributed to a few different optical phenomena combined, the most prominent being reflective diffraction. Using common household items and with a budget of 50 dollars, we attempt to confirm the theory that the collection of ridges on the individual scales on the wing act as transmission gratings, as well as reflective. These results are quantified by the calculation of the ridge separation and comparing this distance to those observed by a scanning electron microscope photo. I. INTRODUCTION In studying the optical property of iridescence, three distinct mechanisms must be discussed. Thin film inter- ference, structured coloring, and reflective diffraction all contribute to the iridescent qualities of a surface. Thin film interference occurs when light strikes a film, some of it enters the film, and the rest is reflected off. The light transmitted through reflects off the bottom of the film and exits the film to interfere with the light that FIG. 1: Up-Close View of Scales from an Optical Microscope originally reflected off the surface of the film. This inter- ference pattern causes a spectrum of color to be visible from white light. Examples of thin film interference in- clude oil slicks and soap bubbles. Structural coloring oc- curs when the structure of the object itself (with various reflective surfaces) produces an interference resulting in vibrant colors. -

The Case of Deirocheline Turtles
bioRxiv preprint doi: https://doi.org/10.1101/556670; this version posted February 21, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Body coloration and mechanisms of colour production in Archelosauria: 2 The case of deirocheline turtles 3 Jindřich Brejcha1,2*†, José Vicente Bataller3, Zuzana Bosáková4, Jan Geryk5, 4 Martina Havlíková4, Karel Kleisner1, Petr Maršík6, Enrique Font7 5 1 Department of Philosophy and History of Science, Faculty of Science, Charles University, Viničná 7, Prague 6 2, 128 00, Czech Republic 7 2 Department of Zoology, Natural History Museum, National Museum, Václavské nám. 68, Prague 1, 110 00, 8 Czech Republic 9 3 Centro de Conservación de Especies Dulceacuícolas de la Comunidad Valenciana. VAERSA-Generalitat 10 Valenciana, El Palmar, València, 46012, Spain. 11 4 Department of Analytical Chemistry, Faculty of Science, Charles University, Hlavova 8, Prague 2, 128 43, 12 Czech Republic 13 5 Department of Biology and Medical Genetics, 2nd Faculty of Medicine, Charles University and University 14 Hospital Motol, V Úvalu 84, 150 06 Prague, Czech Republic 15 6 Department of Food Science, Faculty of Agrobiology, Food, and Natural Resources, Czech University of Life 16 Sciences, Kamýcká 129, Prague 6, 165 00, Czech Republic 17 7 Ethology Lab, Cavanilles Institute of Biodiversity and Evolutionary Biology, University of Valencia, C/ 18 Catedrátic José Beltrán Martinez 2, Paterna, València, 46980, Spain 19 Keywords: Chelonia, Trachemys scripta, Pseudemys concinna, nanostructure, pigments, chromatophores 20 21 Abstract 22 Animal body coloration is a complex trait resulting from the interplay of multiple colour-producing mechanisms. -

Skin Patterning in Octopus Vulgaris and Its Importance for Camouflage
Skin patterning in Octopus vulgaris anditsimportance for camouflage DORSAL ANTERiOR POSTERIOR VENTRAL Wilma Meijer-Kuiper D490 Dqo "Scriptie"entitled: j Skin patterning in Octopus vulgaris and its importance for camouflage Author: Wilma Meijer-Kuiper. Supervisor: Dr. P.J. Geerlink. April, 1993. Cover figure from Wells, 1978. Department of Marine Biology, University of Groningen, Kerklaan 30, 9750 AA Haren, The Netherlands. TABLE OF CONTENTS PAGE Abstract 1 1. Introduction 2 1.1. Camouflage in marine invertebrates 2 1 .2. Cephalopods 2 2. General biology of the octopods 4 2.1. Octopus vulgar/s 4 3. Skin patterning in Octopus 6 3.1. Patterns 6 3.2. Components 8 3.3. Units 9 3.4. Elements 10 3.4.1. Chromatophores 10 3.4.2. Reflector cells and iridophores 16 3.4.3. Leucophores 18 4. The chromatic unit 20 5. Nervous system 22 5.1. Motor units 22 5.2. Electrical stimulation 22 5.3. Projection of nerves on the skin surface 24 5.4. The nervous control system 25 5.5. Neurotransmitters 27 6. Vision in cephalopods, particularly in O.vu!garis 28 6.1. Electro-physiological experiments 28 6.2. Photopigments and receptor cells 28 6.3. Behavioural (training) experiments 29 6.4. Eye movements and optomotor responses 29 7. Camouflage in a colour-blind animal 31 7.1. Matching methods 31 7.2. Countershading reflex 35 8. Conclusive remarks 36 Literature 38 ABSTRACT Camouflage is a method by which animals obtain concealment from other animals by blending in with their environment. Of the cephalopods, the octopods have an extra-ordinary ability to match their surroundings by changing the colour and texture of their skin. -

Iridescence in Cooked Venison – an Optical Phenomenon
Journal of Nutritional Health & Food Engineering Research Article Open Access Iridescence in cooked venison – an optical phenomenon Abstract Volume 8 Issue 2 - 2018 Iridescence in single myofibers from roast venison resembled multilayer interference in having multiple spectral peaks that were easily visible under water. The relationship HJ Swatland of iridescence to light scattering in roast venison was explored using the weighted- University of Guelph, Canada ordinate method of colorimetry. In iridescent myofibers, a reflectance ratio (400/700 nm) showing wavelength-dependent light scattering was correlated with HJ Swatland, Designation Professor CIE (Commission International de l’Éclairage) Y%, a measure of overall paleness Correspondence: Emeritus, University of Guelph, 33 Robinson Ave, Guelph, (r=0.48, P< 0.01). Hence, meat iridescence is an optical phenomenon. The underlying Ontario N1H 2Y8, Canada, Tel 519-821-7513, mechanism, subsurface multilayer interference, may be important for meat colorimetry. Email [email protected] venison, iridescence, interference, reflectance, meat color Keywords: Received: August 23, 2017 | Published: March 14, 2018 Introduction balanced pixel hues that have tricked your eyes to appear white; and when we perceive interference colors, complex interference spectra Iridescence is an enigmatic aspect of meat color with some practical trick our eyes again. As the order of interference increases, the colors 1–4 importance for consumers concerned about green colors in meat. appear to change from metallic -

Octopus Consciousness: the Role of Perceptual Richness
Review Octopus Consciousness: The Role of Perceptual Richness Jennifer Mather Department of Psychology, University of Lethbridge, Lethbridge, AB T1K 3M4, Canada; [email protected] Abstract: It is always difficult to even advance possible dimensions of consciousness, but Birch et al., 2020 have suggested four possible dimensions and this review discusses the first, perceptual richness, with relation to octopuses. They advance acuity, bandwidth, and categorization power as possible components. It is first necessary to realize that sensory richness does not automatically lead to perceptual richness and this capacity may not be accessed by consciousness. Octopuses do not discriminate light wavelength frequency (color) but rather its plane of polarization, a dimension that we do not understand. Their eyes are laterally placed on the head, leading to monocular vision and head movements that give a sequential rather than simultaneous view of items, possibly consciously planned. Details of control of the rich sensorimotor system of the arms, with 3/5 of the neurons of the nervous system, may normally not be accessed to the brain and thus to consciousness. The chromatophore-based skin appearance system is likely open loop, and not available to the octopus’ vision. Conversely, in a laboratory situation that is not ecologically valid for the octopus, learning about shapes and extents of visual figures was extensive and flexible, likely consciously planned. Similarly, octopuses’ local place in and navigation around space can be guided by light polarization plane and visual landmark location and is learned and monitored. The complex array of chemical cues delivered by water and on surfaces does not fit neatly into the components above and has barely been tested but might easily be described as perceptually rich. -
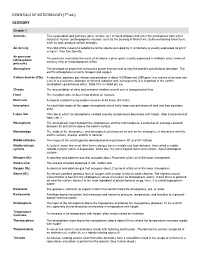
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

Pigment Pattern Formation in Zebrafish 443
MICROSCOPY RESEARCH AND TECHNIQUE 58:442–455 (2002) Pigment Pattern Formation in Zebrafish: A Model for Developmental Genetics and the Evolution of Form 1 1,2 IAN K. QUIGLEY AND DAVID M. PARICHY * 1Section of Integrative Biology, University of Texas at Austin, Austin, Texas 78712 2Section of Molecular, Cell and Developmental Biology and Institute for Cellular and Molecular Biology University of Texas at Austin, Austin, Texas 78712 ABSTRACT The zebrafish Danio rerio is an emerging model organism for understanding vertebrate development and genetics. One trait of both historical and recent interest is the pattern formed by neural crest–derived pigment cells, or chromatophores, which include black melano- phores, yellow xanthophores, and iridescent iridophores. In zebrafish, an embryonic and early larval pigment pattern consists of several stripes of melanophores and iridophores, whereas xanthophores are scattered widely over the flank. During metamorphosis, however, this pattern is transformed into that of the adult, which comprises several dark stripes of melanophores and iridophores that alternate with light stripes of xanthophores and iridophores. In this review, we place zebrafish relative to other model and non-model species; we review what is known about the processes of chromatophore specification, differentiation, and morphogenesis during the develop- ment of embryonic and adult pigment patterns, and we address how future studies of zebrafish will likely aid our understanding of human disease and the evolution of form. Microsc. Res. Tech. 58: 442–455, 2002. © 2002 Wiley-Liss, Inc. INTRODUCTION number of genetic mutants have been isolated that To generate the adult body plan, cells must differen- affect pigmentation. For example, a variety of pigment tiate, proliferate, die, and migrate. -

The Biology and Ecology of the Common Cuttlefish (Sepia Officinalis)
Supporting Sustainable Sepia Stocks Report 1: The biology and ecology of the common cuttlefish (Sepia officinalis) Daniel Davies Kathryn Nelson Sussex IFCA 2018 Contents Summary ................................................................................................................................................. 2 Acknowledgements ................................................................................................................................. 2 Introduction ............................................................................................................................................ 3 Biology ..................................................................................................................................................... 3 Physical description ............................................................................................................................ 3 Locomotion and respiration ................................................................................................................ 4 Vision ................................................................................................................................................... 4 Chromatophores ................................................................................................................................. 5 Colour patterns ................................................................................................................................... 5 Ink sac and funnel organ -

Flip Your Lid for Squid!
FLIP YOUR LID FOR SQUID! Adapted from http://njseagrant.org/wp-content/uploads/2014/03/squid_dissection.pdf PURPOSE Students learn the basics of squid anatomy and physiology by conducting a squid dissection. MATERIALS • frozen whole squid—not “clean” and not cut (get whole squid in frozen section of the grocery store) • hand lenses • dissection pan (plate) • scissors • tweezers • probe or toothpick • paper towels PROCEDURE Dissection of External Anatomy • Have students observe their squids for a few moments looking at the relationship between head and the rest of the body. Then ask students why they think squid are in the family of cephalopods (head-foots). • Tentacles and Arms: Have students count the number of arms. They may use the toothpicks to spread out the squid arms. Ask if all the arms look the same? There should be eight short arms and 2 long arms. Explain that the long thin arms with suckers only at the ends are called tentacles. The tentacles large ends with suckers are known as clubs. The tentacles are used to strike out and capture prey. The eight arms are used to hold onto prey when captured and bring food into its mouth. • Suction Cups: Student may use the magnifying lenses to get a closer look at the suction cups on each arm and tentacles. The suction cups have a small-toothed ring around each one and they are on short stalks. The suction cups are like suckers that help the arms hold onto the prey when it is captured and tries to escape. • Beak and Buccal Mass: Located within the circle of arms students can locate the squids mouth which is a beak. -

Cephalopod Coloration Model. I. Squid Chromatophores and Iridophores
588 J. Opt. Soc. Am. A/Vol. 25, No. 3/March 2008 Sutherland et al. Cephalopod coloration model. I. Squid chromatophores and iridophores Richard L. Sutherland,1,2,* Lydia M. Mäthger,3 Roger T. Hanlon,3 Augustine M. Urbas,1 and Morley O. Stone1 1Air Force Research Laboratory, 3005 Hobson Way, Wright-Patterson Air Force Base, Ohio 45433-7707, USA 2Science Applications International Corporation, 4031 Colonel Glenn Highway, Dayton, Ohio 45431, USA 3Marine Resources Center, Marine Biological Laboratory, 7 MBL Street, Woods Hole, Massachusetts 02543, USA *Corresponding author: [email protected] Received September 11, 2007; revised November 28, 2007; accepted December 3, 2007; posted December 21, 2007 (Doc. ID 87405); published February 7, 2008 We have developed a mathematical model of skin coloration in cephalopods, a class of aquatic animals. Cepha- lopods utilize neurological and physiological control of various skin components to achieve active camouflage and communication. Specific physical processes of this coloration are identified and modeled, utilizing avail- able biological materials data, to simulate active spectral changes in pigment-bearing organs and structural iridescent cells. Excellent agreement with in vitro measurements of squid skin is obtained. A detailed under- standing of the physical principles underlying cephalopod coloration is expected to yield insights into the be- havioral ecology of these animals. © 2008 Optical Society of America OCIS codes: 000.1430, 050.7330, 160.1435, 240.0310, 260.3160, 330.1690. 1. INTRODUCTION -

Rainbow Peacock Spiders Inspire Miniature Super-Iridescent Optics
ARTICLE DOI: 10.1038/s41467-017-02451-x OPEN Rainbow peacock spiders inspire miniature super- iridescent optics Bor-Kai Hsiung 1,8, Radwanul Hasan Siddique 2, Doekele G. Stavenga 3, Jürgen C. Otto4, Michael C. Allen5, Ying Liu6, Yong-Feng Lu 6, Dimitri D. Deheyn 5, Matthew D. Shawkey 1,7 & Todd A. Blackledge1 Colour produced by wavelength-dependent light scattering is a key component of visual communication in nature and acts particularly strongly in visual signalling by structurally- 1234567890 coloured animals during courtship. Two miniature peacock spiders (Maratus robinsoni and M. chrysomelas) court females using tiny structured scales (~ 40 × 10 μm2) that reflect the full visual spectrum. Using TEM and optical modelling, we show that the spiders’ scales have 2D nanogratings on microscale 3D convex surfaces with at least twice the resolving power of a conventional 2D diffraction grating of the same period. Whereas the long optical path lengths required for light-dispersive components to resolve individual wavelengths constrain current spectrometers to bulky sizes, our nano-3D printed prototypes demonstrate that the design principle of the peacock spiders’ scales could inspire novel, miniature light-dispersive components. 1 Department of Biology and Integrated Bioscience Program, The University of Akron, Akron, OH 44325, USA. 2 Department of Medical Engineering, California Institute of Technology, Pasadena, CA 91125, USA. 3 Department of Computational Physics, University of Groningen, 9747 AG Groningen, The Netherlands. 4 19 Grevillea Avenue, St. Ives, NSW 2075, Australia. 5 Scripps Institution of Oceanography (SIO), University of California, San Diego, La Jolla, CA 92093, USA. 6 Department of Electrical and Computer Engineering, University of Nebraska-Lincoln, Lincoln, NE 68588, USA. -

Iridescent Color: from Nature to the Painter's Palette
Downloaded from http://www.mitpressjournals.org/doi/pdf/10.1162/LEON_a_00114 by guest on 25 September 2021 Technical ar T i c l e ce, Tech N e Iridescent Color: From Nature I , Sc to the Painter’s Palette T a b s T r a c T o: Ar nan The shifting rainbow hues of Franziska Schenk iridescence have, until recently, remained exclusive to nature. and Andrew Parker Now, the latest advances in nanotechnology enable the introduction of novel, bio- inspired color-shifting flakes into painting—thereby affording artists potential access to the full spectacle of iridescence. Unfortunately, existing rules rtists have never captured a color as dazzling selves of two distinct types: those of easel painting do not apply A that are stable attributes of material to the new medium; but, as and dynamic as the metallic blue of the Morpho butterfly, which nature inspired the technol- is visible for up to a quarter of a mile. Now, with rapid ad- substances, and those that are “acci- dental,” such as the evanescent col- ogy, an exploration of natural vances in nanoscience and technology, we are beginning to ors of the rainbow and the colors of phenomena can best inform unravel nature’s ingenious manipulation of the flow of light. some birds’ feathers, which change how to overcome this hurdle. Scientific research into natural nanoscale architectures, ca- according to the viewpoint of the Thus, by adopting a biomimetic pable of producing eye-catching optical effects, has led to the spectator [2]. approach, this paper outlines the optical principles underly- development of an ever-expanding range of comparable syn- ing iridescence and provides thetic structures.