Vicarious Reward Response Is Positively Correlated with Functional Connectivity in the Resting State Default Mode Network
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Netease Cloud Music and Big Hit Entertainment Team up to Launch BTS' Song Catalog
NetEase Cloud Music and Big Hit Entertainment team up to launch BTS' song catalog HANGZHOU, China, May 4, 2018 /PRNewswire/ -- NetEase Cloud Music, China's leading music platform, entered into a partnership agreement with South Korea-based Big Hit Entertainment, whereby the Chinese platform will launch BTS' repertoire of songs on the platform. Big Hit Entertainment, a South Korea-based entertainment company focused on music, represents and manages BTS, the company's leading all-male group, which debuted in 2013. BTS is comprised of seven male vocalists: RM, Jin, SUGA, j-hope, Jimin, V and Jung Kook. BTS, the abbreviation for Bangtan Sonyeondan, also stands for Bulletproof Boys and Beyond The Scene. Since the group's first appearance, they have received several major awards at music festivals held in South Korea as well as some international events. In 2017, they were named Top Social Artist by Billboard Music Awards, garnering the group worldwide attention. This February, they were again listed as an award candidate, competing with international superstars. Recently, NetEase Cloud Music launched BTS' widely popular catalog of songs on its platform, including MIC Drop (Steve Aoki Remix) (Feat. Desiigner), DNA, Spring Day, Not Today, Blood Sweat & Tears, Fire, Dope and I need U. In addition to music copyright, NetEase Cloud Music will cooperate with Big Hit Entertainment on artist and music promotion, marking the start of a close collaboration that will cover all aspects of the music business. The two parties look forward to jointly driving the popularity of music artists and their works via multiple channels, while offering higher quality music services to wider audiences. -

The Globalization of K-Pop: the Interplay of External and Internal Forces
THE GLOBALIZATION OF K-POP: THE INTERPLAY OF EXTERNAL AND INTERNAL FORCES Master Thesis presented by Hiu Yan Kong Furtwangen University MBA WS14/16 Matriculation Number 249536 May, 2016 Sworn Statement I hereby solemnly declare on my oath that the work presented has been carried out by me alone without any form of illicit assistance. All sources used have been fully quoted. (Signature, Date) Abstract This thesis aims to provide a comprehensive and systematic analysis about the growing popularity of Korean pop music (K-pop) worldwide in recent years. On one hand, the international expansion of K-pop can be understood as a result of the strategic planning and business execution that are created and carried out by the entertainment agencies. On the other hand, external circumstances such as the rise of social media also create a wide array of opportunities for K-pop to broaden its global appeal. The research explores the ways how the interplay between external circumstances and organizational strategies has jointly contributed to the global circulation of K-pop. The research starts with providing a general descriptive overview of K-pop. Following that, quantitative methods are applied to measure and assess the international recognition and global spread of K-pop. Next, a systematic approach is used to identify and analyze factors and forces that have important influences and implications on K-pop’s globalization. The analysis is carried out based on three levels of business environment which are macro, operating, and internal level. PEST analysis is applied to identify critical macro-environmental factors including political, economic, socio-cultural, and technological. -

Reconciling Pcard Transactions in BOA Works, the Guide
Reconciling PCard Transactions in BOA Works, the Guide Table of Contents RECONCILIATION REPORT ..........................................................................................................................................2 BASIC NAVIGATION IN WORKS ...................................................................................................................................3 • Log In/Lock Out Issues in Works ................................................................................................................3 • Home Page ................................................................................................................................................4 • Action Items...............................................................................................................................................5 • Accounts Dashboard ..................................................................................................................................6 • View Auth Log (Authorization Log)/Check Balance ...................................................................................6 • View Full Details of Account ......................................................................................................................7 • Activate a Card ..........................................................................................................................................9 • Signoff status of transactions ................................................................................................................ -

Between Hybridity and Hegemony in K-Pop's Global Popularity
International Journal of Communication 11(2017), 2367–2386 1932–8036/20170005 Between Hybridity and Hegemony in K-Pop’s Global Popularity: A Case of Girls’ Generation’s American Debut GOOYONG KIM1 Cheyney University of Pennsylvania, USA Examining the sociocultural implications of Korean popular music (K-pop) idol group Girls’ Generation’s (SNSD’s) debut on Late Show With David Letterman, this article discusses how the debut warrants a critical examination on K-pop’s global popularity. Investigating critically how the current literature on K-pop’s success focuses on cultural hybridity, this article maintains that SNSD’s debut clarifies how K-pop’s hybridity does not mean dialectical interactions between American form and Korean content. Furthermore, this article argues that cultural hegemony as a constitutive result of sociohistorical and politico- economic arrangements provides a better heuristic tool, and K-pop should be understood as a part of the hegemony of American pop and neoliberalism. Keywords: Korean popular music, cultural hybridity, cultural hegemony, neoliberalism As one of the most sought-after Korean popular music (K-pop) groups, Girls’ Generation’s (SNSD’s) January 2012 debut on two major network television talk shows in the United States warrants critical reconsideration of the current discourse on cultural hybridity as the basis of K-pop’s global popularity. Prior to Psy’s “Gangnam Style” phenomenon, SNSD’s “The Boys” was the first time a Korean group appeared on an American talk show. It marks a new stage in K-pop’s global reach and influence. With a surge of other K- pop idols gaining global fame, especially in Japan, China, and other Asian countries, SNSD’s U.S. -

Rolex Women's World Golf Rankings Monday, September 05, 2011
Rolex Women's World Golf Rankings Monday, September 05, 2011 Rank Change Name Country Events Total Average 1 -- Yani Tseng TPE 50 778.75 15.57 2 -- Suzann Pettersen NOR 38 464.40 12.22 3 -- Cristie Kerr USA 42 486.25 11.58 4 +1 Na Yeon Choi KOR 51 467.98 9.18 5 -1 Jiyai Shin KOR 56 503.97 9.00 6 -- Sun Ju Ahn KOR 48 387.11 8.06 7 -- Ai Miyazato JPN 51 385.41 7.56 8 -- I.K. Kim KOR 44 325.95 7.41 9 -- Paula Creamer USA 37 271.02 7.32 10 -- Brittany Lincicome USA 45 322.24 7.16 11 -- Stacy Lewis USA 51 359.05 7.04 12 -- Michelle Wie USA 39 252.97 6.49 13 -- Karrie Webb AUS 43 270.87 6.30 14 -- Inbee Park KOR 59 355.17 6.02 15 -- Morgan Pressel USA 46 274.76 5.97 16 -- Chie Arimura JPN 66 370.12 5.61 17 -- Song-Hee Kim KOR 47 251.51 5.35 18 -- Angela Stanford USA 44 234.70 5.33 19 -- Maria Hjorth SWE 45 233.26 5.18 20 -- Sakura Yokomine JPN 65 327.51 5.04 21 -- Miki Saiki JPN 64 310.27 4.85 22 -- Mika Miyazato JPN 52 251.11 4.83 23 -- Ji-Hee Lee KOR 55 256.36 4.66 24 -- Amy Yang KOR 46 212.89 4.63 25 -- So-Yeon Ryu KOR 49 221.41 4.52 26 -- Hee Kyung Seo KOR 46 204.47 4.44 27 -- Yuri Fudoh JPN 49 209.83 4.28 28 -- Anna Nordqvist SWE 47 180.72 3.85 29 -- Mi-Jeong Jeon KOR 60 230.22 3.84 30 -- Catriona Matthew SCO 39 141.85 3.64 31 -- Brittany Lang USA 45 162.93 3.62 32 -- Sun Young Yoo KOR 47 165.53 3.52 33 -- Katherine Hull AUS 53 186.02 3.51 34 +1 Shanshan Feng CHN 40 133.83 3.35 35 -1 Yukari Baba JPN 66 220.52 3.34 36 -- Se-Ri Pak KOR 36 117.97 3.28 37 +3 Na Ri Kim KOR 53 168.14 3.17 38 -1 Sophie Gustafson SWE 51 159.19 3.12 39 +9 Caroline -
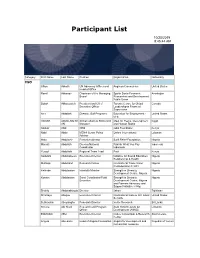
Participant List
Participant List 10/20/2019 8:45:44 AM Category First Name Last Name Position Organization Nationality CSO Jillian Abballe UN Advocacy Officer and Anglican Communion United States Head of Office Ramil Abbasov Chariman of the Managing Spektr Socio-Economic Azerbaijan Board Researches and Development Public Union Babak Abbaszadeh President and Chief Toronto Centre for Global Canada Executive Officer Leadership in Financial Supervision Amr Abdallah Director, Gulf Programs Educaiton for Employment - United States EFE HAGAR ABDELRAHM African affairs & SDGs Unit Maat for Peace, Development Egypt AN Manager and Human Rights Abukar Abdi CEO Juba Foundation Kenya Nabil Abdo MENA Senior Policy Oxfam International Lebanon Advisor Mala Abdulaziz Executive director Swift Relief Foundation Nigeria Maryati Abdullah Director/National Publish What You Pay Indonesia Coordinator Indonesia Yussuf Abdullahi Regional Team Lead Pact Kenya Abdulahi Abdulraheem Executive Director Initiative for Sound Education Nigeria Relationship & Health Muttaqa Abdulra'uf Research Fellow International Trade Union Nigeria Confederation (ITUC) Kehinde Abdulsalam Interfaith Minister Strength in Diversity Nigeria Development Centre, Nigeria Kassim Abdulsalam Zonal Coordinator/Field Strength in Diversity Nigeria Executive Development Centre, Nigeria and Farmers Advocacy and Support Initiative in Nig Shahlo Abdunabizoda Director Jahon Tajikistan Shontaye Abegaz Executive Director International Insitute for Human United States Security Subhashini Abeysinghe Research Director Verite -

Past, Present and Future of Hallyu (Korean Wave)
American International Journal of Contemporary Research Vol. 5, No. 5; October 2015 Past, Present and Future of Hallyu (Korean Wave) Kim Bok-rae Professor Andong national University I. Introduction Hallyu refers to the phenomenon of Korean popular culture which came into vogue in Southeast Asia and mainland China in late 1990s. Especially, hallyu is very popular among young people enchanted with Korean music (K-pop), dramas (K-drama), movies, fashion, food, and beauty in China, Taiwan, Hong Kong, and Vietnam, etc. This cultural phenomenon has been closely connected with multi-layered transnational movements of people, information and capital flows in East Asia. Since the 15th century, East and West have been the two subjects of cultural phenomena. Such East–West dichotomy was articulated by Westerners in the scholarly tradition known as “Orientalism.”During the Age of Exploration (1400–1600), West didn’t only take control of East by military force, but also created a new concept of East/Orient, as Edward Said analyzed it expertly in his masterpiece Orientalism in 1978. Throughout the history of imperialism for nearly 4-5 centuries, west was a cognitive subject, but East was an object being recognized by the former. Accordingly, “civilization and modernization” became the exclusive properties of which West had copyright (?!), whereas East was a “sub-subject” to borrow or even plagiarize from Western standards. In this sense, (making) modern history in East Asia was a compulsive imitation of Western civilization or a catch-up with the West in other wards. Thus, it is interesting to note that East Asian people, after gaining economic power through “compressed modernization,”1 are eager to be main agents of their cultural activities in and through the enjoyment of East Asian popular culture in a postmodern era. -

Global Conservation Translocation Perspectives: 2021. Case Studies from Around the Globe
Global conservation Global conservation translocation perspectives: 2021 translocation perspectives: 2021 IUCN SSC Conservation Translocation Specialist Group Global conservation translocation perspectives: 2021 Case studies from around the globe Edited by Pritpal S. Soorae IUCN SSC Conservation Translocation Specialist Group (CTSG) i The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN or any of the funding organizations concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN. IUCN is pleased to acknowledge the support of its Framework Partners who provide core funding: Ministry of Foreign Affairs of Denmark; Ministry for Foreign Affairs of Finland; Government of France and the French Development Agency (AFD); the Ministry of Environment, Republic of Korea; the Norwegian Agency for Development Cooperation (Norad); the Swedish International Development Cooperation Agency (Sida); the Swiss Agency for Development and Cooperation (SDC) and the United States Department of State. Published by: IUCN SSC Conservation Translocation Specialist Group, Environment Agency - Abu Dhabi & Calgary Zoo, Canada. Copyright: © 2021 IUCN, International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non- commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Citation: Soorae, P. S. -

THE GLOBALIZATION of K-POP by Gyu Tag
DE-NATIONALIZATION AND RE-NATIONALIZATION OF CULTURE: THE GLOBALIZATION OF K-POP by Gyu Tag Lee A Dissertation Submitted to the Graduate Faculty of George Mason University in Partial Fulfillment of The Requirements for the Degree of Doctor of Philosophy Cultural Studies Committee: ___________________________________________ Director ___________________________________________ ___________________________________________ ___________________________________________ Program Director ___________________________________________ Dean, College of Humanities and Social Sciences Date: _____________________________________ Spring Semester 2013 George Mason University Fairfax, VA De-Nationalization and Re-Nationalization of Culture: The Globalization of K-Pop A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at George Mason University By Gyu Tag Lee Master of Arts Seoul National University, 2007 Director: Paul Smith, Professor Department of Cultural Studies Spring Semester 2013 George Mason University Fairfax, VA Copyright 2013 Gyu Tag Lee All Rights Reserved ii DEDICATION This is dedicated to my wife, Eunjoo Lee, my little daughter, Hemin Lee, and my parents, Sung-Sook Choi and Jong-Yeol Lee, who have always been supported me with all their hearts. iii ACKNOWLEDGEMENTS This dissertation cannot be written without a number of people who helped me at the right moment when I needed them. Professors, friends, colleagues, and family all supported me and believed me doing this project. Without them, this dissertation is hardly can be done. Above all, I would like to thank my dissertation committee for their help throughout this process. I owe my deepest gratitude to Dr. Paul Smith. Despite all my immaturity, he has been an excellent director since my first year of the Cultural Studies program. -

Running Head: K-POP FANSHIP, FANDOM, SOCIAL IDENTITY THEORY
Running Head: K-POP FANSHIP, FANDOM, SOCIAL IDENTITY THEORY. 1 Positive Psychosocial Outcomes and Fanship in K-Pop Fans: A Social Identity Theory Perspective. Author Derek A. Laffan Affiliation Dún Laoghaire Institute of Art, Design and Technology Please cite this work as follows: Laffan, D. A. (2020). Positive Psychosocial Outcomes and Fanship in K-Pop Fans: A Social Identity Theory Perspective. Psychological Reports. doi: https://doi.org/10.1177%2F0033294120961524. [Link to article: https://journals.sagepub.com/eprint/ 7RFN3NQNZRFDYWR4C76V/full] Running Head: K-POP FANSHIP, FANDOM, SOCIAL IDENTITY THEORY. 2 Positive Psychosocial Outcomes and Fanship in K-Pop Fans: A Social Identity Theory Perspective. Abstract Korean pop culture (K-Pop) has spread its influence outside of Korea to a worldwide fan audience. The present study investigated the self-categorised K-Pop fandom characteristics that predicted higher levels of K-Pop fanship, and subsequent psychosocial outcomes. Social identity theory was applied as a theoretical framework. In total, 1477 K-Pop fans from 92 predominantly Western countries fully completed an extensive online survey measuring fanship, fandom and psychosocial outcomes (happiness, self-esteem and social connectedness). Results of this study indicated that K- Pop fanship was significantly predicted by a several K-Pop demographic and fandom characteristics. K-Pop fanship was a significant predictor of increased happiness, self-esteem and social connectedness. The study findings advance the application of social identity theory in a K-Pop fan context and the psychological fanship research more broadly. Keywords: Social identity theory, fanship, fandom, K-Pop, self-categorisation. Running Head: K-POP FANSHIP, FANDOM, SOCIAL IDENTITY THEORY. -

Diversity of K-Pop: a Focus on Race, Language, and Musical Genre
DIVERSITY OF K-POP: A FOCUS ON RACE, LANGUAGE, AND MUSICAL GENRE Wonseok Lee A Thesis Submitted to the Graduate College of Bowling Green State University in partial fulfillment of the requirements for the degree of MASTER OF ARTS August 2018 Committee: Jeremy Wallach, Advisor Esther Clinton Kristen Rudisill © 2018 Wonseok Lee All Rights Reserved iii ABSTRACT Jeremy Wallach, Advisor Since the end of the 1990s, Korean popular culture, known as Hallyu, has spread to the world. As the most significant part of Hallyu, Korean popular music, K-pop, captivates global audiences. From a typical K-pop artist, Psy, to a recent sensation of global popular music, BTS, K-pop enthusiasts all around the world prove that K-pop is an ongoing global cultural flow. Despite the fact that the term K-pop explicitly indicates a certain ethnicity and language, as K- pop expanded and became influential to the world, it developed distinct features that did not exist in it before. This thesis examines these distinct features of K-pop focusing on race, language, and musical genre: it reveals how K-pop groups today consist of non-Korean musicians, what makes K-pop groups consisting of all Korean musicians sing in non-Korean languages, what kind of diverse musical genres exists in the K-pop field with two case studies, and what these features mean in terms of the discourse of K-pop today. By looking at the diversity of K-pop, I emphasize that K-pop is not merely a dance- oriented musical genre sung by Koreans in the Korean language. -

SM True Co., Ltd.” to Bring a New Era of Entertainment to Thailand
True Group, in Collaboration with S.M. Entertainment from South Korea, Establishes a Joint-Venture “SM True Co., Ltd.” To Bring A New Era of Entertainment to Thailand Bangkok: August 16, 2011 – True Group’s True Visions Plc, has joined forces with the leading entertainment company from South Korea, S.M. Entertainment Co., Ltd., to co-invest in Thailand’s entertainment industry with the establishment of a joint venture, SM True Co., Ltd. The new company will have exclusive rights to license and distribute all SM products, including CDs and digital music sources as well as merchandise in Thailand. It will also act as the management agency for SM artists in Thailand. This cooperation will bring SM artists from South Korea closer to Thai fans which will drive forward TrueVisions’ entertainment business within True Group. Furthermore, the Thai entertainment business will be able to expand in Asian, offering Thai artists an opportunity to enter the global stage. Attending the opening event of SM True Co., Ltd. are executives from both the investing companies, S.M. Entertainment Co., Ltd. from South Korea represented by Mr. Kim Young-Min, Chief Executive Officer, and Mr. Han Se Min, Director of Strategic Planning Division; and from True Group, led by Mr. Soopakij Chearavanont, Chairman of TrueVisions Plc., Mr. Narong Chearavanont, Director of TrueVisions Plc, Mr. Thiti Nantapatsiri, Managing Director of TrueVisions Plc, and Ms Sasithorn Khunudomsub, Chief Operating Officer of SM True Co., Ltd. Mr. Thiti Nantapatsiri, Managing Director of TrueVisions Plc, said that True Group’s TrueVisions is very honored to have S.M.