Summaries of Wildlife Research Findings 2017
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Evaluating Grassland Wildlife Exposure to Soybean Aphid Insecticides on Public Lands in Minnesota
EVALUATING GRASSLAND WILDLIFE EXPOSURE TO SOYBEAN APHID INSECTICIDES ON PUBLIC LANDS IN MINNESOTA Katelin Goebel1 and Nicole M. Davros SUMMARY OF FINDINGS Increasing evidence suggests that pesticides may be an important factor explaining declines in grassland-dependent wildlife in agricultural landscapes. Minnesota Department of Natural Resource (MNDNR) wildlife managers and members of the public have reported concerns about foliar-application insecticides in particular. Such insecticides are used on a variety of crops but their use has been especially important for controlling soybean aphid outbreaks in Minnesota. Concerns have been raised about the impacts of chlorpyrifos, a broad-spectrum organophosphate, and other foliar-application insecticides on water quality and human health, prompting the Minnesota Department of Agriculture (MDA) to release guidelines for voluntary best management practices for their use. Although lab studies have shown chlorpyrifos and other insecticides used to target aphids are highly toxic to non-target organisms, including economically important game species and pollinators, few studies have investigated the environmentally-relevant exposure of free-ranging wildlife to these chemicals. Our objective was to assess the direct and indirect exposure of grassland wildlife to the 3 most common soybean aphid insecticides (i.e., chlorpyrifos, lambda-cyhalothrin, and bifenthrin) along a gradient from soybean field edge to grassland interior. During summer 2017 and 2018, we sampled 5 treatment and 4 control sites across western and southern Minnesota. We detected chlorpyrifos at all distances examined (0-400 m) at both treatment and control sites, suggesting that some background level of chlorpyrifos exposure is occurring in the environment regardless of landowner activities in the adjacent row crop field. -

SELECTION of Entomopathogenic Nematodes Against Dalaca Pallens (Lepidoptera: Hepialidae)
RESEARCH SELECTION OF EntoMOPathoGENIC NEMatoDES AGAINST DALACA PALLENS (LEPIDOPTERA: HEPIALIDAE) Alexis Maldonado1, Loreto Merino1, and Andrés France1* Dalaca pallens (Blanchard) is a pest with great impact on grassland production in Chile. The objective of this research was to evaluate the use of entomopathogenic nematodes (EPN) against D. pallens larvae. Twenty EPN isolates, collected along Chile, were used for nematode selection. The experimental unit consisted of five 50-mL plastic vials filled withNothofagus dombeyi sawdust and inoculated with 40 dauers per vial. The experimental design was a completely randomized with four replicates per treatment. The results showed that isolate QU N3 (Steinernema australe) and QU N13 (Steinernema unicornum) produced the highest mortality, with 100 and 95% respectively and no differences (P < 0.001) between both. Later, these two isolates were used to calculate the lethal concentration (LC50 and LC90). Concentrations of 0, 10, 20, 30, 40, and 50 dauers per 50-mL plastic vials filled withNothofagus sawdust were evaluated. Then, a single larva of D. pallens was added to each vial and incubated for 13 d. The results showed that LC50 and LC90 were equivalent to 14 and 39 dauers mL-1 for isolate QU N3, while for isolate QUN13 these figures were 14 and 48 dauers mL-1. Consequently, there are native isolates of EPN with the ability to control D. pallens larvae, which require future test under field conditions. Key words: Biological control, Steinernema australe, Steinernema unicornium, lethal concentration. alaca pallens (Blanchard) (Lepidoptera: bacteria, belonging to the genus Xenorhabdus and Hepialidae) is an economically important pest, Photorhabdus, respectively (Emelianoff et al., 2007). -

Entomopathogenic Nematology in Latin America: a Brief History, Current Re- Search and Future Prospects
Accepted Manuscript Entomopathogenic nematology in Latin America: A brief history, current re- search and future prospects Ernesto San-Blas, Raquel Campos-Herrera, Claudia Dolinski, Caio Monteiro, Vanessa Andaló, Luis Garrigós Leite, Mayra G. Rodríguez, Patricia Morales- Montero, Adriana Sáenz-Aponte, Carolina Cedano, Juan Carlos López-Nuñez, Eleodoro Del Valle, Marcelo Doucet, Paola Lax, Patricia D. Navarro, Francisco Báez, Pablo Llumiquinga, Jaime Ruiz-Vega, Abby Guerra-Moreno, S. Patricia Stock PII: S0022-2011(18)30180-0 DOI: https://doi.org/10.1016/j.jip.2019.03.010 Reference: YJIPA 7192 To appear in: Journal of Invertebrate Pathology Received Date: 31 May 2018 Revised Date: 31 December 2018 Accepted Date: 29 March 2019 Please cite this article as: San-Blas, E., Campos-Herrera, R., Dolinski, C., Monteiro, C., Andaló, V., Garrigós Leite, L., Rodríguez, M.G., Morales-Montero, P., Sáenz-Aponte, A., Cedano, C., Carlos López-Nuñez, J., Del Valle, E., Doucet, M., Lax, P., Navarro, P.D., Báez, F., Llumiquinga, P., Ruiz-Vega, J., Guerra-Moreno, A., Patricia Stock, S., Entomopathogenic nematology in Latin America: A brief history, current research and future prospects, Journal of Invertebrate Pathology (2019), doi: https://doi.org/10.1016/j.jip.2019.03.010 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. -

Pest Alert: Brown Marmorated Stink Bug Halyomorpha Halys
OREGON DEPARTMENT OF AGRICULTURE FACT SHEETS AND PEST ALERTS Pest Alert: Brown Marmorated Stink Bug Halyomorpha halys Introduction Brown Marmorated Stink Bug The brown marmorated stink bug (BMSB), Halyomorpha adult nymph newly-hatched halys, is an Asian species first detected in North America nymphs in Pennsylvania in 1996, and in Oregon in 2004. BMSB has since been detected in 43 states. In Oregon it is es- tablished statewide, in the western region from Portland to Ashland, and in the north east to Hood River. More recently it has been found in coastal counties and is likely still expanding its range and increasing in abundance around Oregon. A threat to Oregon agriculture BMSB is a major agricultural pest in Asia, attacking many crops. It is a significant agricultural pest in the Mid-At- lantic states of the U.S., attacking tree fruits, peppers, tomatoes, corn, berries, grapes, soybeans, melons, and even damaging young trees by feeding through the bark. BMSB is known to feed on over 170 species of plants. The insect threatens an estimated $21 billion worth of crops in the United States alone. Some commercial agri- cultural damage by BMSB has been reported in Oregon. Some home gardeners have reported extensive damage to beans, cucumbers, raspberries, hops, and several species A brown marmorated stink bug feeding on a mature hazelnut. BMSB of ornamental plants. is able to feed on tree nuts through the shell using its long mouthparts. Damage to crops Stink bugs feed by inserting their long, straw-like mouth parts into plants and sucking out the liquid inside. -

Boxelder Bug
BOXELDER BUG Integrated Pest Management for Home Gardeners and Landscape Professionals The western boxelder bug (Boisea rubrolineata) is often a nuisance pest around and in homes. Boxelder bugs usually feed on the leaves, flowers, and seedpods of the female or seedbearing box elder tree (Acer negundo), although they may also subsist on male box elder trees and occasionally occur on maple and ash trees. They may feed on the fruits of almond, apple, cherry, peach, Figure 1. Boxelder bug adult and nymphs. Figure 2. Young nymph of western box- pear, and plum trees, and on grapes, (J. K. Clark) elder bug, Boisea rubrolineata. where their feeding punctures cause (J. K. Clark) the fruit to become deformed. Large numbers of the bug usually occur only on female box elder trees. IDENTIFICATION When full grown, the boxelder bug is about 1/2 inch long and one-third as wide. Adults are mostly black and have three red lines on the pronotum of the thorax (one down the middle and on each margin) and several fine Figure 3. Boxelder bug eggs on leaf. Figure 4. Adult squash bug. red lines on each wing (Figure 1). The (J. K. Clark) (J. K. Clark) wings lie flat on the bug’s back when it is at rest. The abdomen is red. The young nymphs are bright red (Figure 2) and when approaching adulthood, become marked with black and begin to develop black wing pads. Eggs are yellow when first laid but become red as nymphs develop inside (Figure 3). Boxelder bugs are true bugs (Order: Hemiptera) in the family Rhopalidae. -

Boxelder Bug Nuisance Management for Homeowners
CIS 1155 Boxelder Bug Nuisance Management for Homeowners by Danielle Gunn and Edward John Bechinski Boxelder bugs are a common nuisance pest in Figure 1. Idaho homes and yards. Although not particu- Comparative larly harmful, these insects can be aggravating life-size boxelder when they are searching for places to spend the bug 1st-stage winter. nymph (left) and adult (right). This publication will help you understand both the seasonal biology of boxelder bugs in Idaho, and landscape features that increase pest prob- lems. We discuss the relative importance of these insects as pests. Practical steps you can take to reduce nuisance problems include alternatives to insecticides and safe, effective insecticide use. Identification Boxelder bugs develop through three life stages: eggs, nymphs, and adults. Figure 1 shows the actual body sizes of a newly hatched nymph and a mature adult. Adults are the most commonly encountered life Figure 2. Adult boxelder bugs, Boisea trivittata, are stage. Adult boxelder bugs are flattened, elon- distinctively marked with red lines on a slate-gray back- gate insects approximately one-half-inch long ground. (not including antennae). Overall upper body color is slate gray to black. Reddish orange lines appear behind the head and along the sides of the body (Figure 2). The rest of the body under the wings is red with two rows of black spots. Legs and antennae are black. Eggs. Small red eggs occur in clusters on box- elder and maple trees. Elongate eggs one-six- teenth-inch long are laid in groups of about ten on the bark and leaves of host trees and sur- rounding areas. -
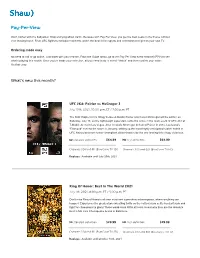
Pay-Per-View
Pay-Per-View Don’t bother with the babysitter. Stop worrying about traffic. Because with Pay Per View, you get the best seats in the house without ever leaving home. From UFC fights to exclusive concerts, watch the best in live sports and entertainment right on your own TV. Ordering made easy No need to call or go online. Just order with your remote. From the Guide menu, go to the Pay Per View event channel (PPV) to see what’s playing this month. Once you’ve made your selection, all you need to do is select “Watch” and then confirm your order. It’s that easy. What’s new this month? UFC 264: Poirier vs McGregor 3 July 10th, 2021, 10:00 p.m. ET / 7:00 p.m. PT The final chapter in the trilogy between Dustin Poirier and Conor McGregor will be written on Saturday, July 10, as the lightweight superstars settle the score in the main event of UFC 264 at T-Mobile Arena in Las Vegas. After Ireland's McGregor defeated Poirier in 2014, Louisiana's "Diamond" evened the score in January, setting up the most highly anticipated rubber match in UFC history between former champions determined to be the one leaving this trilogy victorious. SD standard definition $64.99 HD high definition $64.99 Channels 324 and 611 (BlueCurve TV SD) Channels 300 and 601 (BlueCurve TV HD) Replays: Available until July 25th, 2021 Ring Of Honor: Best In The World 2021 July 11h, 2021, 8:00 p.m. ET / 5:00 p.m. -

SIDE EFFECTS of FUNGICIDES USED in CUCURBITACEOUS CROP on Trichogramma Atopovirilia OATMAN & PLATNER (HYMENOPTERA: TRICHOGRAMATIDAE)
SCIENTIFIC NOTE 323 SIDE EFFECTS OF FUNGICIDES USED IN CUCURBITACEOUS CROP ON Trichogramma atopovirilia OATMAN & PLATNER (HYMENOPTERA: TRICHOGRAMATIDAE) Dirceu Pratissoli1, André M. Milanez1, Wagner F. Barbosa2*, Flávio N. Celestino1, Gilberto S. Andrade3, and Ricardo A. Polanczyk1 ABSTRACT Trichogramma spp. (Hymenoptera: Trichogrammatidae) can control Diaphania hyalinata Linnaeus (Lepidoptera: Pyralidae). On the other hand, pesticides may reduce the efficiency of natural enemies. The objective was to evaluate the side-effects of fungicides used in the production of cucurbitaceous crops on Trichogramma atopovirilia Oatman & Platner parasitizing D. hyalinata eggs. The fungicides used in bioassays were: azoxystrobin (0.08 g active ingredient [ai] L-1), chlorothalonil (2.00 g ai L-1), mancozeb (1.60 g ai L-1), tebuconazole (0.25 g ai L-1) and thiophanate-methyl (0.49 g ai L-1). Cardboards with 30 D. hyalinata eggs previously immersed in fungicide solutions and distilled water (control) were offered separately to 20 newly emerged T. atopovirilia females in glass tubes. Parasitism, parasitism reduction, emergence, sex ratio, and number of individuals per egg were evaluated. The fungicides chlorothalonil, thiophanate-methyl and tebuconazole reduced parasitism of T. atopovirilia by 43.37, 27.64 and 18.51%, respectively. However, parasitism with azoxystrobin (79.21%) was higher than the control (67.37%) (P ≤ 0.05). Chlorothalonil, thiophanate-methyl and tebuconazole reduced emergence by 73.77, 75.62 and 79.35% (P ≤ 0.05), respectively. Azoxystrobin and thiophanate-methyl reduced the sex ratio by 0.77 and 0.76 (P ≤ 0.05), respectively. Fungicides did not reduce the number of individuals per egg. The fungicides azoxystrobin and mancozeb were selective for T. -

That Are N O Ttuurito
THAT AREN O US009802899B2TTUURITO ( 12) United States Patent (10 ) Patent No. : US 9 ,802 , 899 B2 Heilmann et al. ( 45 ) Date of Patent: Oct . 31, 2017 ( 54 ) HETEROCYCLIC COMPOUNDS AS CO7D 401/ 12 ( 2006 .01 ) PESTICIDES C07D 403 /04 (2006 .01 ) CO7D 405 / 12 (2006 . 01) (71 ) Applicant : BAYER CROPSCIENCE AG , C07D 409 / 12 ( 2006 .01 ) Monheim (DE ) C070 417 / 12 (2006 . 01) (72 ) Inventors: Eike Kevin Heilmann , Duesseldorf AOIN 43 /60 ( 2006 .01 ) (DE ) ; Joerg Greul , Leverkusen (DE ) ; AOIN 43 /653 (2006 . 01 ) Axel Trautwein , Duesseldorf (DE ) ; C07D 249 /06 ( 2006 . 01 ) Hans- Georg Schwarz , Dorsten (DE ) ; (52 ) U . S . CI. Isabelle Adelt , Haan (DE ) ; Roland CPC . .. C07D 231/ 40 (2013 . 01 ) ; AOIN 43 / 56 Andree , Langenfeld (DE ) ; Peter ( 2013 .01 ) ; A01N 43 /58 ( 2013 . 01 ) ; AOIN Luemmen , Idstein (DE ) ; Maike Hink , 43 /60 (2013 .01 ) ; AOIN 43 /647 ( 2013 .01 ) ; Markgroeningen (DE ); Martin AOIN 43 /653 ( 2013 .01 ) ; AOIN 43 / 76 Adamczewski , Cologne (DE ) ; Mark ( 2013 .01 ) ; A01N 43 / 78 ( 2013 .01 ) ; A01N Drewes, Langenfeld ( DE ) ; Angela 43/ 82 ( 2013 .01 ) ; C07D 231/ 06 (2013 . 01 ) ; Becker , Duesseldorf (DE ) ; Arnd C07D 231 /22 ( 2013 .01 ) ; C07D 231/ 52 Voerste , Cologne (DE ) ; Ulrich ( 2013 .01 ) ; C07D 231/ 56 (2013 .01 ) ; C07D Goergens, Ratingen (DE ) ; Kerstin Ilg , 249 /06 (2013 . 01 ) ; C07D 401 /04 ( 2013 .01 ) ; Cologne (DE ) ; Johannes -Rudolf CO7D 401/ 12 ( 2013 . 01) ; C07D 403 / 04 Jansen , Monheim (DE ) ; Daniela Portz , (2013 . 01 ) ; C07D 403 / 12 ( 2013 . 01) ; C07D Vettweiss (DE ) 405 / 12 ( 2013 .01 ) ; C07D 409 / 12 ( 2013 .01 ) ; C07D 417 / 12 ( 2013 .01 ) ( 73 ) Assignee : BAYER CROPSCIENCE AG , (58 ) Field of Classification Search Monheim ( DE ) ??? . -

Hemiptera: Heteroptera) with a Review of the Eggs of This Family
ACTA ENTOMOLOGICA MUSEI NATIONALIS PRAGAE Published 30.vi.2010 Volume 50(1), pp. 75–95 ISSN 0374-1036 The external morphology of eggs of three Rhopalidae species (Hemiptera: Heteroptera) with a review of the eggs of this family Jitka VILÍMOVÁ & Markéta ROHANOVÁ Charles University, Faculty of Science, Department of Zoology, Viničná 7, CZ-128 44 Praha 2, Czech Republic; e-mail: [email protected]; [email protected] Abstract. The external morphology of eggs and manner of oviposition of three rhopalid species, Brachycarenus tigrinus (Schilling, 1829), Chorosoma schillingi (Schilling, 1829) and Rhopalus (Aeschyntelus) maculatus (Fieber, 1837) are described. The eggs were studied using Scanning Electron Microscopy (SEM), and the results complete previous observations.The emphasis of the study is on the characteristics of eggs and details of oviposition in representatives of the family Rhopalidae. The chorionic origin of attachment stalk was confi rmed only in the Chorosomatini. A completely smooth egg chorion was recognized in R. (A.) maculatus, as a unique condition within at least the Pentatomomorpha. Key words. Rhopalidae, Rhopalini, Chorosomatini, Brachycarenus tigrinus, Cho- rosoma schillingi, Rhopalus (Aeschyntelus) maculatus, egg structure, micropylar processes, chorion, attachment stalk, oviposition Introduction Heteroptera eggs have a stable shape due to a sclerotized chorion. Egg morphology is helpful for taxonomic and phylogenetic purposes. The morphology of heteropteran eggs varies distinctly among taxa; for details see two monographs: SOUTHWOOD (1956) and COB- BEN (1968). Both authors mentioned that the eggs of the coreoid family Rhopalidae have a specifi c morphological pattern (e.g. two micropylar processes). COBBEN (1968) not only compared the morphology of heteropteran eggs but made phylogenetic inferences from their important characters. -

Summary of Sexual Abuse Claims in Chapter 11 Cases of Boy Scouts of America
Summary of Sexual Abuse Claims in Chapter 11 Cases of Boy Scouts of America There are approximately 101,135sexual abuse claims filed. Of those claims, the Tort Claimants’ Committee estimates that there are approximately 83,807 unique claims if the amended and superseded and multiple claims filed on account of the same survivor are removed. The summary of sexual abuse claims below uses the set of 83,807 of claim for purposes of claims summary below.1 The Tort Claimants’ Committee has broken down the sexual abuse claims in various categories for the purpose of disclosing where and when the sexual abuse claims arose and the identity of certain of the parties that are implicated in the alleged sexual abuse. Attached hereto as Exhibit 1 is a chart that shows the sexual abuse claims broken down by the year in which they first arose. Please note that there approximately 10,500 claims did not provide a date for when the sexual abuse occurred. As a result, those claims have not been assigned a year in which the abuse first arose. Attached hereto as Exhibit 2 is a chart that shows the claims broken down by the state or jurisdiction in which they arose. Please note there are approximately 7,186 claims that did not provide a location of abuse. Those claims are reflected by YY or ZZ in the codes used to identify the applicable state or jurisdiction. Those claims have not been assigned a state or other jurisdiction. Attached hereto as Exhibit 3 is a chart that shows the claims broken down by the Local Council implicated in the sexual abuse. -

Boxelder Bug Boisea Trivittata (Say); Family: Rhopalidae
IDL INSECT DIAGNOSTIC LABORATORY Cornell University, Dept. of Entomology, 2144 Comstock Hall, Ithaca NY 14853-2601 Boxelder Bug Boisea trivittata (Say); Family: Rhopalidae Boxelder bug (adult); actual size about 1/2 inch long. Photo © 2012 by Jason J. Dombroskie. Injury The boxelder bug may be a pest of outdoor trees as well as a household nuisance. It is the latter that is of most concern to homeowners. The bugs overwinter as adults and nymphs in protected dry places, often in wall voids or attics of houses and buildings. During warm days in the fall and again in the spring, the bugs become active and invade homes, creating a nuisance. They do not feed while indoors. Description The adult boxelder bug is about 1/2 inch in length, and brownish-black in color, with red stripes on the thorax and wing margins. The body is also bright red. Eggs are straw-yellow to rusty-red in color and are not often seen, as they are deposited on boxelder trees (Acer negundo) or other maples, or near the trees. The nymphs, which are found on the trees, are bright red in color with the front half darker. Nymphs resemble adults but do not have fully developed wings and are not able to reproduce. The change from nymph to adult is a gradual one. Life History The boxelder bugs pass the winter in the adult and sometimes nymphal stages in dry, sheltered places where they have accumulated in gregarious masses. They often choose buildings or houses as a protected place to overwinter. When weather warms up in the spring, the bugs leave their places of hibernation to fly to boxelder trees, where they deposit their eggs.