Scathophaga Stercoraria): Investigating Sperm Competition Models by Histological Observation D.J
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Selection on Morphological Traits and Fluctuating Asymmetry by a Fungal Parasite in the Yellow Dung Fly
bioRxiv preprint doi: https://doi.org/10.1101/136325; this version posted September 14, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. 1 2 3 4 5 6 7 Blanckenhorn WU. 2017. 8 Selection on morphological traits and fluctuating asymmetry by a 9 fungal parasite in the yellow dung fly. 10 bioRxiv 136325, http://dx.doi.org/10.1101/136325 11 12 A Preprint reviewed and recommended by Peer Community 13 Evolutionary Biology: 14 http://dx.doi.org/10.24072/pci.evolbiol.100027 15 16 17 bioRxiv preprint doi: https://doi.org/10.1101/136325; this version posted September 14, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. Wolf U. Blanckenhorn Selection by a fungal parasite in dung flies Selection on morphological traits and fluctuating asymmetry by a fungal parasite in the yellow dung fly WOLF U. BLANCKENHORN Department of Evolutionary Biology and Environmental Studies, University of Zürich, Winterthurerstrasse 190, CH-8057 Zürich, Switzerland; Fax: +41 44 635.4780; E-mail: [email protected] Abstract Evidence for selective disadvantages of large body size remains scarce in general. Previous phenomenological studies of the yellow dung fly Scathophaga stercoraria have demonstrated strong positive sexual and fecundity selection on male and female size. -

Repeatability of Sperm Size in Outbred and Inbred Scathophaga Stercoraria (Diptera: Scathophagidae)
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2007 Repeatability of sperm size in outbred and inbred Scathophaga stercoraria (Diptera: Scathophagidae) Bernasconi, G ; Ward, P I ; Hellriegel, B Abstract: Variability in male gametic traits can depend on several genetic and environmental factors such as developmental instability as a consequence of inbreeding, developmental noise during spermatogenesis, or age- or condition-dependent changes in allocation to sperm cells. Variation in sperm size is particularly evident in species that produce more than one sperm morph but also occurs among males in sperm- monomorphic species. Both discrete and continuous sperm size variation have been implicated in male fertilization success when the sperm of several males directly compete for fertilization of the same set of ova. In this study, we investigated among-male variation in sperm length in field-collected, outbred male Scathophaga stercoraria (L.) flies, as well as in flies from the same natural population that hadbeen subjected to 15 and 16 generations of inbreeding under laboratory conditions. Among-male variation in sperm length was significant and repeatable over subsequent matings in both inbred and outbred flies. We conclude that sperm length can be used as an individual male marker in sperm competition studies and that significant repeatability of sperm length supports heritability for this trait DOI: https://doi.org/10.4039/n06-063 Posted at the Zurich Open Repository and Archive, University of Zurich ZORA URL: https://doi.org/10.5167/uzh-154393 Journal Article Published Version Originally published at: Bernasconi, G; Ward, P I; Hellriegel, B (2007). -

A Fungal Parasite Selects Against Body Size but Not Fluctuating Asymmetry in Swiss Subalpine Yellow Dung Flies
Alpine Entomology 5 2021, 27–35 | DOI 10.3897/alpento.5.65653 A fungal parasite selects against body size but not fluctuating asymmetry in Swiss subalpine yellow dung flies Wolf U. Blanckenhorn1 1 Department of Evolutionary Biology and Environmental Studies, University of Zürich, Winterthurerstrasse 190, CH-8057, Zürich, Switzerland http://zoobank.org/0D01A23B-D327-4F92-9726-93AAD929CAFD Corresponding author: Wolf U. Blanckenhorn ([email protected]) Academic editor: Patrick Rohner ♦ Received 8 March 2021 ♦ Accepted 25 May 2021 ♦ Published 11 June 2021 Abstract Evidence for selective disadvantages of large body size remains scarce in general. Previous studies of the yellow dung fly Scatho- phaga stercoraria have demonstrated strong positive sexual and fecundity selection on male and female size. Nevertheless, the body size of flies from a Swiss study population has declined by ~10% 1993–2009. Given substantial heritability of body size, this neg- ative evolutionary response of an evidently positively selected trait suggests important selective factors being missed. An episodic epidemic outbreak of the fungus Entomophthora scatophagae permitted assessment of natural selection exerted by this fatal parasite. Fungal infection varied over the season from ~50% in the cooler and more humid spring and autumn to almost 0% in summer. The probability of dying from fungal infection increased with adult fly body size. Females never laid any eggs after infection, so there was no fungus effect on female fecundity beyond its impact on mortality. Large males showed their typical mating advantage in the field, but this positive sexual selection was nullified by fungal infection. Mean fluctuating asymmetry of paired appendages (legs, wings) did not affect the viability, fecundity or mating success of yellow dung flies in the field. -

Experimental Evidence for Testis Size Evolution Via Sperm Competition
Paper 198 Disc Ecology Letters, (2001) 4 : 10±13 LETTER Experimental evidence for testis size evolution via sperm competition Abstract David J. Hosken and Paul I. Ward Sperm competition theory predicts increased spermatogenic investment with Zoological Museum, University increased sperm competition risk when competition is numerical. There is ample of ZuÈrich, Winterthurerstr 190, correlational evidence for this relationship in a wide range of taxa. However, as with ZuÈrich 8057, Switzerland. E-mail: all correlations, this does not establish cause and effect. Nevertheless, there are no [email protected] published experimental studies of the evolutionary influence of sperm competition on testis size. We report here on evolutionary responses of testis size to variation in sperm competition intensity in the yellow dung fly. Experimental flies were divided across two treatments, polyandrous or monogamous, with four replicates of each. There was a rapid evolutionary response in testis size resulting from selection via sperm competition, with larger testes found when sperm competition intensity was greatest. These results provide direct experimental evidence of evolutionary change consistent with macro-evolutionary patterns found across a wide range of taxa. Keywords Polyandry, Scathophaga, Scatophaga, sperm competition, testes. Ahed Ecology Letters (2001) 4 : 10±13 Bhed Ched Dhed INTRODUCTION Simmons & Parker 1992; Ward 1993, 1998, 2000; Ref marker reviewed in Hosken 1999). Typically, males that copulate Fig marker Sperm competition (Parker 1970a) theory predicts that second (or last) fertilize about 80% of the subsequent Table mar- when competition is fundamentally analogous to a raffle, clutch, at least under common laboratory conditions ker relative sperm number in competition will be the primary (Parker 1970b). -

Dipterists Digest
Dipterists Digest 2018 Vol. 25 No. 2 Cover illustration: Palloptera usta (Meigen, 1826) (Pallopteridae), male, on a rotten birch log at Glen Affric (NH 28012832), 4 November 2018. © Alan Watson Featherstone. In Britain, a predominantly Scottish species, having strong associations with Caledonian pine forest, but also developing in wood of broad-leaved trees. Rearing records from under bark of Betula (3), Fraxinus (1), Picea (18), Pinus (21), Populus (2) and Quercus (1) were cited by G.E. Rotheray and R.M. Lyszkowski (2012. Pallopteridae (Diptera) in Scotland. Dipterists Digest (Second Series ) 19, 189- 203). Apparently a late date, as the date range given by Rotheray and Lyszkowski ( op. cit .) for both adult captures and emergence dates from puparia was 13 May to 29 September. Dipterists Digest Vol. 25 No. 2 Second Series 2018 th Published 27 February 2019 Published by ISSN 0953-7260 Dipterists Digest Editor Peter J. Chandler, 606B Berryfield Lane, Melksham, Wilts SN12 6EL (E-mail: [email protected]) Editorial Panel Graham Rotheray Keith Snow Alan Stubbs Derek Whiteley Phil Withers Dipterists Digest is the journal of the Dipterists Forum . It is intended for amateur, semi- professional and professional field dipterists with interests in British and European flies. All notes and papers submitted to Dipterists Digest are refereed. Articles and notes for publication should be sent to the Editor at the above address, and should be submitted with a current postal and/or e-mail address, which the author agrees will be published with their paper. Articles must not have been accepted for publication elsewhere and should be written in clear and concise English. -

Size-Dependent Energetics of Metamorphosis in the Yellow Dung Fly, Scathophaga Stercoraria
Evolutionary Ecology Research, 2009, 11: 1111–1130 Size-dependent energetics of metamorphosis in the yellow dung fly, Scathophaga stercoraria Constanze Reim, Christian Kaufmann*, and Wolf U. Blanckenhorn Zoologisches Museum, Universität Zürich-Irchel, Zürich, Switzerland ABSTRACT Competing hypotheses: (1) Large body size confers more efficient energy use (relative efficiency hypothesis). (2) Large body size requires more energy to be sustained, a disadvantage when food is limited (absolute energy demand hypothesis). Organism: Yellow dung flies, Scathophaga stercoraria (Diptera: Scathophagidae), artificially selected for large and small body size in the laboratory for 24 generations to augment the available phenotypic body sizes. Methods: Larvae were reared under limited and unlimited food (dung) conditions, and the energy content of pupae was measured at the beginning and the end of the pupal stage in different, size-matched individuals. Conclusions: Over the pupal period, lipids and glycogen decreased whereas sugar content increased. Net energy loss per unit body mass was higher at unlimited food. Contrary to expectation, males (the larger sex) lost less energy than females. Large selection line pupae showed the highest absolute and mass-specific energy loss during metamorphosis, indicating a correlated physiological response to body size selection because phenotypic body sizes do not differ between the lines at limited dung. We conclude that energetic costs due to greater absolute energy demand of larger individuals during the pupal phase outweigh the benefits due to greater metabolic efficiency. Keywords: artificial selection, body size, development, energy reserves, insect pupa, life history, metabolic rate, survival. INTRODUCTION The life history and fitness of most organisms are most crucially influenced by body size. -

Zootaxa 2441: 27–40 (2010) ISSN 1175-5326 (Print Edition) Article ZOOTAXA Copyright © 2010 · Magnolia Press ISSN 1175-5334 (Online Edition)
Zootaxa 2441: 27–40 (2010) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2010 · Magnolia Press ISSN 1175-5334 (online edition) Systematic ambiguity in the well-established model system insect Scathophaga stercoraria (Diptera: Scathophagidae): sister species S. soror revealed by molecular evidence MARCO VALERIO BERNASCONI1*, DAVID BERGER1 & WOLF U. BLANCKENHORN1 1Zoological Museum, Institute of Evolutionary Biology and Environmental Studies, University of Zurich, Winterthurerstrasse 190, CH- 8057 Zurich, Switzerland E-mails: [email protected]; [email protected]; [email protected] *corresponding author Abstract Even for well-established insect model systems, such as the yellow dung fly Scathophaga stercoraria (Linnaeus) (Diptera: Scathophagidae), there may be hidden systematic ambiguities that require clarification. Dung flies from the Afrotropical Region have been considered (i) as con-specific and not different from all the other Holarctic Scathophaga stercoraria; (ii) as a local and peculiar African subspecies of S. stercoraria (Scathophaga stercoraria soror Wiedemann), or (iii) as a separate valid species (Scathophaga soror Wiedemann). Our study represents an attempt, based on mitochondrial (COI, 12S, and 16S), nuclear (ITS2) as well as microsatellite markers, to clarify this problem. Results strongly suggest that S. soror is a separate taxon from S. stercoraria. Due to the importance of S. stercoraria as a model system for studies in ecology, behaviour and evolution, -

OECD/OCDE 228 Adopted: 29 July 2016
OECD/OCDE 228 Adopted: 29 July 2016 OECD GUIDELINEFOR THE TESTING OF CHEMICALS Determination of Developmental Toxicity to Dipteran Dung Flies(Scathophaga stercoraria L. (Scathophagidae), Musca autumnalis De Geer (Muscidae)) INTRODUCTION 1. This Test Guideline is designed to assess the effects of the test chemical on the development of dung dwelling life stages of dung-dependent dipteran species. In this test, insects are exposed under controlled conditions [1] to the test chemical spiked into the dung. An extended test, in which the flies are exposed to dung originating from livestock treated with the test substance, is described in ANNEX 4. 2. Scathophaga stercoraria L. (Scathophagidae) and Musca autumnalis De Geer (Muscidae)) are considered to be suitable indicator species for estimating the developmental toxicity of parasiticides on dung dependant Diptera for the following main reasons: Collectively, the species cover a wide geographic range. S. stercoraria and M. autumnalis are widespread in Europe, Asia, Africa and North America [2, 3, 4, 5, 6, 7]. By including a Musca-species the area which is covered by this guideline is extended: For example, the Australian species M. vetustissima could be tested in the same way as M. autumnalis, while S. stercoraria is not occurring there. Despite some overlap, the two species differ in their temperature preferences: S. stercoraria avoids temperatures >25° and has been found in Iceland and at high altitudes in the Alps while M. autumnalis prefers just the opposite. 3. Both species are dung-dependent, are multi-voltine, do not undergo obligate diapause and are easy to culture and have a short life-cycle which makes it possible to determine effects on development and survival in the laboratory. -

Unlocking the “Black Box”: Internal Female Genitalia in Sepsidae (Diptera) Evolve Fast and Are Species-Specific
Puniamoorthy et al. BMC Evolutionary Biology 2010, 10:275 http://www.biomedcentral.com/1471-2148/10/275 RESEARCH ARTICLE Open Access Unlocking the “Black box”: internal female genitalia in Sepsidae (Diptera) evolve fast and are species-specific Nalini Puniamoorthy1,2*†, Marion Kotrba3†, Rudolf Meier2† Abstract Background: The species-specificity of male genitalia has been well documented in many insect groups and sexual selection has been proposed as the evolutionary force driving the often rapid, morphological divergence. The internal female genitalia, in sharp contrast, remain poorly studied. Here, we present the first comparative study of the internal reproductive system of Sepsidae. We test the species-specificity of the female genitalia by comparing recently diverged sister taxa. We also compare the rate of change in female morphological characters with the rate of fast-evolving, molecular and behavioral characters. Results: We describe the ectodermal parts of the female reproductive tract for 41 species representing 21 of the 37 described genera and define 19 morphological characters with discontinuous variation found in eight structures that are part of the reproductive tract. Using a well-resolved molecular phylogeny based on 10 genes, we reconstruct the evolution of these characters across the family [120 steps; Consistency Index (CI): 0.41]. Two structures, in particular, evolve faster than the rest. The first is the ventral receptacle, which is a secondary sperm storage organ. It accounts for more than half of all the evolutionary changes observed (7 characters; 61 steps; CI: 0.46). It is morphologically diverse across genera, can be bi-lobed or multi-chambered (up to 80 chambers), and is strongly sclerotized in one clade. -

Fitness Consequences of the Combined Effects of Veterinary and Agricultural Pesticides on a Non-Target Insect Hayat Mahdjoub1, W
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2020 Fitness consequences of the combined effects of veterinary and agricultural pesticides on a non-target insect Mahdjoub, Hayat ; Blanckenhorn, Wolf U ; Lüpold, Stefan ; Roy, Jeannine ; Gourgoulianni, Natalia ; Khelifa, Rassim Abstract: Pesticides and veterinary products that are globally used in farming against pests and parasites are known to impact non-target beneficial organisms. While most studies have tested the lethal and sub-lethal effects of single chemicals, species are exposed to multiple contaminants that might interact and exacerbate the toxic responses of life-history fitness components. Here we experimentally tested an ecotoxicological scenario that is likely to be widespread in nature, with non-target dung communities being exposed both to cattle parasiticides during the larval stage and to agricultural insecticides during their adult life. We assessed the independent and combined consumptive effects of varying ivermectin and spinosad concentration on juvenile life-history and adult reproductive traits of the widespread yellow dung fly (Scathophaga stercoraria; Diptera: Scathophagidae). Larval exposure to ivermectin prolonged development time and reduced egg-to-adult survival, body size, and the magnitude of the male-biased sexual size dimorphism. The consumption by the predatory adult flies of spinosad-contaminated prey showed an additional, independent (from ivermectin) negative effect on female clutch size, and subsequent egg hatching success, but not on the body size and sexual size dimorphism of their surviving offspring. However, there were interactive synergistic effects of both contaminants on offspring emergence and body size. Our results document adverse effects of the combination of different chemicals on fitness components of a dung insect, highlighting transgenerational effects of adult exposure to contaminants for their offspring. -
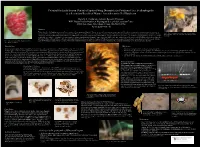
Potential for Early Season Control of Spotted Wing Drosophila by Predatory Flies, Scathophagidae As a Secondary Benefit of Manure Amendments in Red Raspberry
Potential for Early Season Control of Spotted Wing Drosophila by Predatory Flies, Scathophagidae as a Secondary Benefit of Manure Amendments in Red Raspberry Beverly S. Gerdeman, Assistant Research Professor WSU Mount Vernon Northwest Washington Research & Extension Center 16650 State Route 536, Mount Vernon, WA 98273-4768 [email protected] Abstract Yellow dung flies, Scathophaga stercoraria (L.) are members of Scathophagidae (Diptera). They are predatory, feeding mainly on other dipterans. They breed in dung and are abundant around livestock and in Fig. 2. Dung flies exhibit sexual dimorphism, males are husky, agricultural fields where dung is utilized as a soil amendment. The appearance of large numbers of dung flies in a red raspberry field in early spring 2015, provoked an investigation into their potential to feed on hairy and golden, while females are svelte, striped and blend with spotted wing drosophila, Drosophila suzukii (Matsumura), SWD, the #1 pest of red raspberries and resident in Washington State. Dissections verified dung fly mouthparts are equipped for predation. Cage studies the manure. provided forensic evidence, decapitated and empty drosophila cadavers, proof that dung flies would capture and feed on SWD. Molecular analyses detected the presence of SWD DNA in fresh collected dung flies, suggesting SWD is now a part of their regular diet. Integration of dung flies into a conventional red raspberry SWD control program remains possible if spring manure applications are timed to coincide with the Fig. 1. Spotted wing drosophila, Drosophila suzukii, is return of SWD founder females while meeting the 90-day (NOP Standard) PHI for berries not touching the ground, or if pathogen-free biodigester by-products can be tweaked to attract dung flies. -

Hered 401 Master..Hered 401 .. Page655
Heredity 80 (1998) 655–659 Received 29 January 1998 Short Review Intraspecific variation in sperm size characters PAUL I. WARD* Zoologisches Museum der Universit¨at Z¨urich, Winterthurerstr. 190, 8057 Z¨urich, Switzerland Interspecific variation in quantitative characters of sperm is evolution. Size differences between the sperm of competing considerable and can sometimes be related to aspects of males could be used in controlled matings to investigate the sperm competition. In contrast, continuous intraspecific vari- fates of sperm within female tracts and storage organs, but ation is often ignored; for example, many studies concentrate care should be taken that size characters may not be neutral on measuring a species-typical sperm size. However, statis- markers. Furthermore, the genetic determination of sperm tically significant variation amongst males in a variety of size is considerable (h2 = 0.56–0.92 for total length and a sperm size characters, particularly total length and a number number of head characters) and determinants are at least of head characters, has been reported in at least 13 species. sometimes sex-linked, with the result that particular care Furthermore, sperm size has been shown to influence the must go into experimental design if genetic effects are to be outcomes of processes involved in sperm competition in two fully elucidated. Studies combining genetical and behavioural species, with larger sperm being more successful in both effects of intraspecific variation in sperm morphology should cases. It is suggested here that intraspecific variation may be be particularly rewarding. a widespread phenomena, further examination of which Keywords: fertilization, heritability, heterospermic insemi- could contribute considerably to our understanding of sperm nation, sex linkage, sperm competition, sperm storage.