University of Florida Thesis Or Dissertation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Winter Precipitation Liquid–Ice Phase Transitions Revealed with Polarimetric Radar and 2DVD Observations in Central Oklahoma
MAY 2017 B U K O V CICETAL. 1345 Winter Precipitation Liquid–Ice Phase Transitions Revealed with Polarimetric Radar and 2DVD Observations in Central Oklahoma PETAR BUKOVCIC ´ NOAA/National Severe Storms Laboratory, and Cooperative Institute for Mesoscale Meteorological Studies, and School of Meteorology, and Advanced Radar Research Center, University of Oklahoma, Norman, Oklahoma DUSAN ZRNIC´ NOAA/National Severe Storms Laboratory, Norman, Oklahoma GUIFU ZHANG School of Meteorology, and Advanced Radar Research Center, University of Oklahoma, Norman, Oklahoma (Manuscript received 30 June 2016, in final form 8 November 2016) ABSTRACT Observations and analysis of an ice–liquid phase precipitation event, collected with an S-band polarimetric KOUN radar and a two-dimensional video disdrometer (2DVD) in central Oklahoma on 20 January 2007, are presented. Using the disdrometer measurements, precipitation is classified either as ice pellets or rain/freezing rain. The disdrometer observations showed fast-falling and slow-falling particles of similar size. The vast majority (.99%) were fast falling with observed velocities close to those of raindrops with similar sizes. In contrast to the smaller particles (,1 mm in diameter), bigger ice pellets (.1.5 mm) were relatively easy to distinguish because their shapes differ from the raindrops. The ice pellets were challenging to detect by looking at conventional polarimetric radar data because of the localized and patchy nature of the ice phase and their occurrence close to the ground. Previously published findings referred to cases in which ice pellet areas were centered on the radar location and showed a ringlike structure of enhanced differential reflectivity ZDR and reduced copolar correlation coefficient rhv and horizontal reflectivity ZH in PPI images. -

Downloaded 10/01/21 08:52 PM UTC 186 WEATHER and FORECASTING VOLUME 16
FEBRUARY 2001 NOTES AND CORRESPONDENCE 185 Further Investigation of a Physically Based, Nondimensional Parameter for Discriminating between Locations of Freezing Rain and Ice Pellets ROBERT M. RAUBER,LARRY S. OLTHOFF, AND MOHAN K. RAMAMURTHY Department of Atmospheric Sciences, University of Illinois at Urbana±Champaign, Urbana, Illinois KENNETH E. KUNKEL Midwestern Climate Center, Illinois State Water Survey, Champaign, Illinois 9 December 1999 and 16 August 2000 ABSTRACT The general applicability of an isonomogram developed by Czys and coauthors to diagnose the position of the geographic boundary between freezing precipitation (freezing rain or freezing drizzle) and ice pellets (sleet or snow grains) was tested using a 25-yr sounding database consisting of 1051 soundings, 581 where stations were reporting freezing drizzle, 391 reporting freezing rain, and 79 reporting ice pellets. Of the 1051 soundings, only 306 clearly had an environmental temperature and moisture pro®le corresponding to that assumed for the isonomogram. This pro®le consisted of a three-layer atmosphere with 1) a cold cloud layer aloft that is a source of ice particles, 2) a midlevel layer where the temperature exceeds 08C and ice particles melt, and 3) a surface layer where T , 08C. The remaining soundings did not conform to the pro®le either because 1) the freezing precipitation was associated with the warm rain process or 2) the ice pellets formed due to riming rather than melting and refreezing. For soundings conforming to the pro®le, the isonomogram showed little diagnostic skill. Freezing rain or freezing drizzle occurred about 50% of the time that ice pellets were expected. -

Ice Nucleators, Bacterial Cells and Pseudomonas
1 Ice nucleators, bacterial cells and Pseudomonas syringae in 2 precipitation at Jungfraujoch 3 Emiliano Stopelli1, Franz Conen1, Caroline Guilbaud2, Jakob Zopfi3, Christine Alewell1, 4 Cindy E. Morris2 5 1Environmental Geosciences, University of Basel, 4056 Basel, Switzerland 6 2INRA PACA, UR 0407 Plant Pathology Research Unit, 84143 Montfavet, France 7 3Acquatic and Stable Isotope Biogeochemistry, University of Basel, 4056 Basel, Switzerland 8 Correspondence to: Emiliano Stopelli ([email protected]) and Franz Conen ([email protected]) 9 10 Abstract Ice nucleation is a means by which the deposition of an airborne microorganism can be accelerated 11 under favourable meteorological conditions. Analysis of 56 snow samples collected at the high altitude 12 observatory Jungfraujoch (3580 m a.s.l.) revealed an order of magnitude larger dynamic range of ice nucleating 13 particles active at -8 °C (INPs-8) compared to the total number of bacterial cells (of which 60 % was on average 14 alive). This indicates a shorter atmospheric residence time for INPs-8. Furthermore, concentrations of INPs-8 15 decreased much faster, with an increasing fraction of water precipitated from the air mass prior to sampling, than -1 16 the number of total bacterial cells. Nevertheless, at high wind speeds (> 50 km h ) the ratio of INPs-8 to total 17 bacterial cells largely remained in a range between 10-2 to 10-3, independent of prior precipitation, likely because 18 of recent injections of particles in regions upwind. Based on our field observations, we conclude that ice 19 nucleators travel shorter legs of distance with the atmospheric water cycle than the majority of bacterial cells. -

Hydrologic Highways Between Microbial Communities of the Plant Microbiome? John Tolan Van Stan, Cindy E
Precipitation partitioning - Hydrologic highways between microbial communities of the plant microbiome? John Tolan van Stan, Cindy E. Morris, K Aung, Y Kuzyakov, D Magyar, E A Rebollar, M Remus-Emsermann, Stéphane Uroz, Philippe Vandenkoornhuyse To cite this version: John Tolan van Stan, Cindy E. Morris, K Aung, Y Kuzyakov, D Magyar, et al.. Precipitation partitioning - Hydrologic highways between microbial communities of the plant microbiome?. van Stan John Toland; Gutmann Ethan; Friesen Jan. Precipitation partitioning by vegetation, Springer Nature, pp.229-251, 2020, 978-3-030-29701-5. 10.1007/978-3-030-29702-2_14. hal-02389738 HAL Id: hal-02389738 https://hal.archives-ouvertes.fr/hal-02389738 Submitted on 26 Mar 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Chapter 14. Precipitation partitioning— Hydrologic highways between microbial communities of the plant microbiome? John T. Van Stan II1*, Cindy E. Morris2, Kyaw Aung3, Yakov Kuzyakov4, Donát Magyar5, Eria A. Rebollar6, Mitja Remus-Emsermann7, Stéphane Uroz8, Philippe Vandenkoornhuyse9 -

News and Notes
346 BULLETIN AMERICAN METEOROLOGICAL SOCIETY NEWS AND NOTES Air Force Scientists Develop Technique The feasibility of dissipating supercooled clouds by for Cutting Holes in "Cold" Clouds seeding with dry-ice was recognized many years ago. Airborne equipment, previously developed to do this, A new technique for creating holes in supercooled cloud crushed large blocks of dry-ice into particles suitable for layers has been developed by Air Force Cambridge Re- seeding purposes. search Laboratories scientists. During recent flight tests This equipment had two particular limitations. First, using this technique, holes more than 3 miles wide were these machines produced about half powdered dry-ice created in supercooled clouds. which evaporated immediately upon dispersal from the Major James F. Church, project scientist in AFCRL's aircraft and was wasted. Second, the logistical problem Meteorological Research Laboratory is directing research of supplying enough dry-ice where needed and the high on the dissipation of supercooled stratiform clouds. The evaporational losses suffered during storage until used, program is under the technical management of the Air severely limited the utility of this technique. Force System Command's Electronic Systems Division. The Cloudbuster makes ice pellets by passing liquid The principal objective of this research is to provide C02 through an expansion nozzle and then compacting Air Force cargo and transport pilots with an economical the solid dry-ice powder, which is then formed, into proper capability to seed supercooled clouds with dry-ice pellets, size. The Cloudbuster has been designed with the capa- made on the aircraft and dispersed at will, to create bility of making different-sized pellets at the touch of a sizable holes in such cloud decks. -

Survival and Ice Nucleation Activity of Pseudomonas Syringae Strains Exposed To
bioRxiv preprint doi: https://doi.org/10.1101/408906; this version posted September 6, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 Survival and ice nucleation activity of Pseudomonas syringae strains exposed to 2 simulated high-altitude atmospheric conditions 3 Gabriel Guarany de Araujoa, Fabio Rodriguesb, Fabio Luiz Teixeira Gonçalvesc, Douglas 4 Galanted,* 5 a Interunities Graduate Program in Biotechnology, University of São Paulo, Brazil 6 b Department of Fundamental Chemistry, Institute of Chemistry, University of São Paulo, 7 Brazil 8 c Department of Atmospheric Sciences, Institute of Astronomy, Geophysics and 9 Atmospheric Sciences, University of São Paulo, Brazil 10 d Brazilian Synchrotron Light Laboratory, Brazilian Center for Research in Energy and 11 Materials, Brazil 12 * Corresponding author. Brazilian Synchrotron Light Laboratory, Av. Giuseppe Máximo 13 Scolfaro, 10000, CEP 13083‐100, Campinas, São Paulo, Brazil, Tel.: +551935175081; Fax: 14 +551935121004. E-mail address: [email protected] 15 Running title: Survival of ice nucleating bacteria on the atmosphere 16 ABSTRACT 17 The epiphytic bacterium Pseudomonas syringae produces the most efficient and 18 well-studied biological ice nuclei (IN) known. Bioaerosols containing these cells have been 19 proposed to influence cloud glaciation, an important process in the initiation of 20 precipitation. The presence of this species has been reported on rain, snow, and cloud water 1 bioRxiv preprint doi: https://doi.org/10.1101/408906; this version posted September 6, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. -

METAR/SPECI Reporting Changes for Snow Pellets (GS) and Hail (GR)
U.S. DEPARTMENT OF TRANSPORTATION N JO 7900.11 NOTICE FEDERAL AVIATION ADMINISTRATION Effective Date: Air Traffic Organization Policy September 1, 2018 Cancellation Date: September 1, 2019 SUBJ: METAR/SPECI Reporting Changes for Snow Pellets (GS) and Hail (GR) 1. Purpose of this Notice. This Notice coincides with a revision to the Federal Meteorological Handbook (FMH-1) that was effective on November 30, 2017. The Office of the Federal Coordinator for Meteorological Services and Supporting Research (OFCM) approved the changes to the reporting requirements of small hail and snow pellets in weather observations (METAR/SPECI) to assist commercial operators in deicing operations. 2. Audience. This order applies to all FAA and FAA-contract weather observers, Limited Aviation Weather Reporting Stations (LAWRS) personnel, and Non-Federal Observation (NF- OBS) Program personnel. 3. Where can I Find This Notice? This order is available on the FAA Web site at http://faa.gov/air_traffic/publications and http://employees.faa.gov/tools_resources/orders_notices/. 4. Cancellation. This notice will be cancelled with the publication of the next available change to FAA Order 7900.5D. 5. Procedures/Responsibilities/Action. This Notice amends the following paragraphs and tables in FAA Order 7900.5. Table 3-2: Remarks Section of Observation Remarks Section of Observation Element Paragraph Brief Description METAR SPECI Volcanic eruptions must be reported whenever first noted. Pre-eruption activity must not be reported. (Use Volcanic Eruptions 14.20 X X PIREPs to report pre-eruption activity.) Encode volcanic eruptions as described in Chapter 14. Distribution: Electronic 1 Initiated By: AJT-2 09/01/2018 N JO 7900.11 Remarks Section of Observation Element Paragraph Brief Description METAR SPECI Whenever tornadoes, funnel clouds, or waterspouts begin, are in progress, end, or disappear from sight, the event should be described directly after the "RMK" element. -

Classification of Three Airborne Bacteria and Proposal Of
International Journal of Systematic and Evolutionary Microbiology (2002), 52, 445–456 DOI: 10.1099/ijs.0.01682-0 Classification of three airborne bacteria and proposal of Hymenobacter aerophilus sp. nov. 1 Institut fu$ r Bakteriologie, Sandra Buczolits,1,2 Ewald B. M. Denner,2 Dietmar Vybiral,2 Mykologie und Hygiene, 1,2 3 1,2 Veterina$ rmedizinische Monika Wieser, Peter Ka$ mpfer and Hans-Ju$ rgen Busse Universita$ t, Veterina$ rplatz 1, A-1210 Wien, Austria 2 Author for correspondence: Hans-Ju$ rgen Busse. Tel: j43 1 25077 2128. Fax: j43 1 25077 2190. Institut fu$ r Mikrobiologie e-mail: Hans-Juergen.Busse!vu-wien.ac.at und Genetik, Universita$ t Wien, A-1030 Wien, Austria Three aerobic, Gram-negative, rod-shaped, non-spore-forming, red-pigmented, 3 Institut fu$ r Angewandte airborne bacteria (I/26-Cor1T, I/32A-Cor1 and I/74-Cor2) collected in the Museo Mikrobiologie, Justus- Liebig-Universita$ t Giessen, Correr (Venice, Italy) were investigated to determine their taxonomic status by D-35390 Giessen, Germany analysing their biochemical, physiological and chemotaxonomic features and the GMC content of genomic DNA and by comparing their genomic fingerprints. Additionally, the almost complete 16S rRNA gene sequence of strain I/26-Cor1T was analysed. The three strains were nearly identical in their morphological, physiological, biochemical and chemotaxonomic properties. The strains contained a menaquinone system with the predominant menaquinone MK-7 and a fatty acid profile with C15:0 anteiso, C15:0 iso and C16:1 predominant. Phosphatidylethanolamine and several unidentified lipids were detected in the polar lipid profiles. The polyamine pattern consisted of sym-homospermidine as the major compound. -
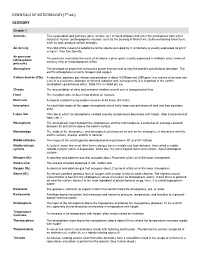
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

Taxonomy JN869023
Species that differentiate periods of high vs. low species richness in unattached communities Species Taxonomy JN869023 Bacteria; Actinobacteria; Actinobacteria; Actinomycetales; ACK-M1 JN674641 Bacteria; Bacteroidetes; [Saprospirae]; [Saprospirales]; Chitinophagaceae; Sediminibacterium JN869030 Bacteria; Actinobacteria; Actinobacteria; Actinomycetales; ACK-M1 U51104 Bacteria; Proteobacteria; Betaproteobacteria; Burkholderiales; Comamonadaceae; Limnohabitans JN868812 Bacteria; Proteobacteria; Betaproteobacteria; Burkholderiales; Comamonadaceae JN391888 Bacteria; Planctomycetes; Planctomycetia; Planctomycetales; Planctomycetaceae; Planctomyces HM856408 Bacteria; Planctomycetes; Phycisphaerae; Phycisphaerales GQ347385 Bacteria; Verrucomicrobia; [Methylacidiphilae]; Methylacidiphilales; LD19 GU305856 Bacteria; Proteobacteria; Alphaproteobacteria; Rickettsiales; Pelagibacteraceae GQ340302 Bacteria; Actinobacteria; Actinobacteria; Actinomycetales JN869125 Bacteria; Proteobacteria; Betaproteobacteria; Burkholderiales; Comamonadaceae New.ReferenceOTU470 Bacteria; Cyanobacteria; ML635J-21 JN679119 Bacteria; Proteobacteria; Betaproteobacteria; Burkholderiales; Comamonadaceae HM141858 Bacteria; Acidobacteria; Holophagae; Holophagales; Holophagaceae; Geothrix FQ659340 Bacteria; Verrucomicrobia; [Pedosphaerae]; [Pedosphaerales]; auto67_4W AY133074 Bacteria; Elusimicrobia; Elusimicrobia; Elusimicrobiales FJ800541 Bacteria; Verrucomicrobia; [Pedosphaerae]; [Pedosphaerales]; R4-41B JQ346769 Bacteria; Acidobacteria; [Chloracidobacteria]; RB41; Ellin6075 -

Ancestry and Adaptive Radiation of Bacteroidetes As Assessed by Comparative Genomics
1 Ancestry and adaptive radiation of Bacteroidetes as assessed by comparative genomics 2 3 Raul Munoza,b,*, Hanno Teelinga, Rudolf Amanna and Ramon Rosselló-Mórab,* 4 5 a Department of Molecular Ecology, Max Planck Institute for Marine Microbiology, D-28359 6 Bremen, Germany. 7 b Marine Microbiology Group, Department of Ecology and Marine Resources, Institut Mediterrani 8 d’Estudis Avançats (CSIC-UIB), E-07190 Esporles, Balearic Islands, Spain. 9 10 * Corresponding authors: 11 Raul Munoz, Marine Microbiology Group, Carrer Miquel Marquès 21, 07190 Esporles, Illes 12 Balears, Spain. e-mail: [email protected] 13 Ramon Rosselló-Móra, Marine Microbiology Group, Carrer Miquel Marquès 21, 07190 Esporles, 14 Illes Balears, Spain. e-mail: [email protected] 15 16 17 18 Keywords + 19 Bacteroidetes, Na -NQR, alternative complex III, caa3 cytochrome oxidase, gliding, T9SS. 20 21 Abbreviations 22 m.s.i.: median sequence identity. 23 24 25 26 27 ABSTRACT 28 As of this writing, the phylum Bacteroidetes comprises more than 1,500 described species with 29 diverse ecological roles. However, there is little understanding of archetypal Bacteroidetes traits on 30 a genomic level. We compiled a representative set of 89 Bacteroidetes genomes and used pairwise 31 reciprocal best match gene comparisons and gene syntenies to identify common traits that allow to 32 trace Bacteroidetes’ evolution and adaptive radiation. Highly conserved among all studied 33 Bacteroidetes was the type IX secretion system (T9SS). Class-level comparisons furthermore 34 suggested that the ACIII-caa3COX super-complex evolved in the ancestral aerobic bacteroidetal 35 lineage, and was secondarily lost in extant anaerobic Bacteroidetes. -

Bioprecipitation Studies, 1921-1927. Springfield, IL
STATE OF ILLINOIS DEPARTMENT OF REGISTRATION AND EDUCATION DIVISION OF THE STATE WATER SURVEY A. M. BUSWELL, Chief BULLETIN NO. 25 BIOPRECIPITATION STUDIES 1921-1927 BY A. M. BUSWELL, R. A. SHIVE, & S. L. NEAVE [Printed by authority of the State of Illinois] URBANA, ILLINOIS ORGANIZATION STATE OF ILLINOIS LEN SMALL, Governor DEPARTMENT OF REGISTRATION AND EDUCATION A. M. SHELTON, Director Board of Natural Resources and Conservation Advisers A. M. SHELTON, Chairman WILLIAM A. NOYES, Chemistry HENRY C. COWLES, Forestry. Secretary. WILLIAM TRELEASE, Biology. JOHN W. ALVORD, Engineering. C. M. THOMPSON, Representing . EDSON S. BASTIN, Geology. the President of the University of Illinois. State Water Survey Division Committee A. M. SHELTON WILLIAM A. NOYES C. M. THOMPSON JOHN W. ALVORD STATE WATER SURVEY DIVISION A. M. BUSWELL, Chief JEFFERSONS PRINTING AND STATIONERY COMPANY SPRINGFIELO. ILLINOIS 105 1500 19ZB 2 CONTENTS Page Letter of transmittal.................................................................................4 Introduction and summary, by A. M. BUSWELL 5 Part I—Aeration, by A. M. BUSWELL and S. L. NEAVE 22 Bubble aeration 24 Mechanical aeration 27 Application to the activated—sludge process 31 Summary 33 Appendix to Part I—B. O. D. experiment with small nidus tanks 35 Part II—The role of bioprecipitation in sewage treatment, by R. A. SHIVE with A. M. BUSWELL 37 Proof of bioprecipitation 39 Construction of experimental sewage treatment plant 47 Operation and control of plant.................................................55 Data on operation of nidus tank.... .. 57 Sludge filtering experiments 80 Trickling filter experiments 80 Summary of results of nidus tank operation 81 Part III—Recent experiments with nidus tank, by A.