Two Examples of Animal Like Protists
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Morphological and Phylogenetic Study of the Genus Chondria (Rhodomelaceae, Rhodophyta)
Title A morphological and phylogenetic study of the genus Chondria (Rhodomelaceae, Rhodophyta) Author(s) Sutti, Suttikarn Citation 北海道大学. 博士(理学) 甲第13264号 Issue Date 2018-06-29 DOI 10.14943/doctoral.k13264 Doc URL http://hdl.handle.net/2115/71176 Type theses (doctoral) File Information Suttikarn_Sutti.pdf Instructions for use Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP A morphological and phylogenetic study of the genus Chondria (Rhodomelaceae, Rhodophyta) 【紅藻ヤナギノリ属(フジマツモ科)の形態学的および系統学的研究】 Suttikarn Sutti Department of Natural History Sciences, Graduate School of Science Hokkaido University June 2018 1 CONTENTS Abstract…………………………………………………………………………………….2 Acknowledgement………………………………………………………………………….5 General Introduction………………………………………………………………………..7 Chapter 1. Morphology and molecular phylogeny of the genus Chondria based on Japanese specimens……………………………………………………………………….14 Introduction Materials and Methods Results and Discussions Chapter 2. Neochondria gen. nov., a segregate of Chondria including N. ammophila sp. nov. and N. nidifica comb. nov………………………………………………………...39 Introduction Materials and Methods Results Discussions Conclusion Chapter 3. Yanagi nori—the Japanese Chondria dasyphylla including a new species and a probable new record of Chondria from Japan………………………………………51 Introduction Materials and Methods Results Discussions Conclusion References………………………………………………………………………………...66 Tables and Figures 2 ABSTRACT The red algal tribe Chondrieae F. Schmitz & Falkenberg (Rhodomelaceae, Rhodophyta) currently -

Biology and Systematics of Heterokont and Haptophyte Algae1
American Journal of Botany 91(10): 1508±1522. 2004. BIOLOGY AND SYSTEMATICS OF HETEROKONT AND HAPTOPHYTE ALGAE1 ROBERT A. ANDERSEN Bigelow Laboratory for Ocean Sciences, P.O. Box 475, West Boothbay Harbor, Maine 04575 USA In this paper, I review what is currently known of phylogenetic relationships of heterokont and haptophyte algae. Heterokont algae are a monophyletic group that is classi®ed into 17 classes and represents a diverse group of marine, freshwater, and terrestrial algae. Classes are distinguished by morphology, chloroplast pigments, ultrastructural features, and gene sequence data. Electron microscopy and molecular biology have contributed signi®cantly to our understanding of their evolutionary relationships, but even today class relationships are poorly understood. Haptophyte algae are a second monophyletic group that consists of two classes of predominately marine phytoplankton. The closest relatives of the haptophytes are currently unknown, but recent evidence indicates they may be part of a large assemblage (chromalveolates) that includes heterokont algae and other stramenopiles, alveolates, and cryptophytes. Heter- okont and haptophyte algae are important primary producers in aquatic habitats, and they are probably the primary carbon source for petroleum products (crude oil, natural gas). Key words: chromalveolate; chromist; chromophyte; ¯agella; phylogeny; stramenopile; tree of life. Heterokont algae are a monophyletic group that includes all (Phaeophyceae) by Linnaeus (1753), and shortly thereafter, photosynthetic organisms with tripartite tubular hairs on the microscopic chrysophytes (currently 5 Oikomonas, Anthophy- mature ¯agellum (discussed later; also see Wetherbee et al., sa) were described by MuÈller (1773, 1786). The history of 1988, for de®nitions of mature and immature ¯agella), as well heterokont algae was recently discussed in detail (Andersen, as some nonphotosynthetic relatives and some that have sec- 2004), and four distinct periods were identi®ed. -

Ultrastructural and Transcriptome Changes of Free-Living Sporangial Filaments in Pyropia Yezoensis Affected by Light and Culture
Ultrastructural and transcriptome changes of free-living sporangial filaments in Pyropia yezoensis affected by light and culture density Bangxiang He1, Xiujun Xie1, and Guangce Wang1 1Institute of Oceanology, Chinese Academy of Sciences April 28, 2020 Abstract In the life cycle of Pyropia yezoensis, sporangial filaments connect conchocelis and thallus, but the mechanisms of maturation and conchospore release of sporangial filaments are poorly understood. We found that the morphological change from vegetative growth form (hollow cells) to reproductive form (bipartite cells), and the release of conchospores from bipartite cells were all closely correlated with culture density and light intensity. Bipartite cells formed at low density (50{1,000 fragments/mL) and when stimulated by high light levels (40{100 mmol photons m-2 s-1), but conchospore release was inhibited at such light intensities. At high densities (5,000{10,000 fragments/mL), sporangial filaments retained the hollow cell morphology and rarely formed bipartite cells. Ultrastructural observation showed that the degradation of autophagosome-like structures in vacuoles caused the typical hollow form. Transcriptome analysis indicated that adaptive responses to environmental changes, mainly autophagy, endocytosis and phosphatidylinositol metabolism, caused the morphological transformation of free-living sporangial filaments. Meanwhile, the extensive promotion of energy accumulation under high light levels promoted vegetative growth of sporangial filaments, and thus inhibited conchospore release from bipartite cells. These results provide a theoretical basis for maturation of sporangial filaments and release of conchospores in P. yezoensis and other related species. Main text Ultrastructural and transcriptome changes of free-living sporangial filaments in Pyropia yezoensis affected by light and culture density Abstract In the life cycle of Pyropia yezoensis , sporangial filaments connect conchocelis and thallus, but the mecha- nisms of maturation and conchospore release of sporangial filaments are poorly understood. -

Predatory Flagellates – the New Recently Discovered Deep Branches of the Eukaryotic Tree and Their Evolutionary and Ecological Significance
Protistology 14 (1), 15–22 (2020) Protistology Predatory flagellates – the new recently discovered deep branches of the eukaryotic tree and their evolutionary and ecological significance Denis V. Tikhonenkov Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences, Borok, 152742, Russia | Submitted March 20, 2020 | Accepted April 6, 2020 | Summary Predatory protists are poorly studied, although they are often representing important deep-branching evolutionary lineages and new eukaryotic supergroups. This short review/opinion paper is inspired by the recent discoveries of various predatory flagellates, which form sister groups of the giant eukaryotic clusters on phylogenetic trees, and illustrate an ancestral state of one or another supergroup of eukaryotes. Here we discuss their evolutionary and ecological relevance and show that the study of such protists may be essential in addressing previously puzzling evolutionary problems, such as the origin of multicellular animals, the plastid spread trajectory, origins of photosynthesis and parasitism, evolution of mitochondrial genomes. Key words: evolution of eukaryotes, heterotrophic flagellates, mitochondrial genome, origin of animals, photosynthesis, predatory protists, tree of life Predatory flagellates and diversity of eu- of the hidden diversity of protists (Moon-van der karyotes Staay et al., 2000; López-García et al., 2001; Edg- comb et al., 2002; Massana et al., 2004; Richards The well-studied multicellular animals, plants and Bass, 2005; Tarbe et al., 2011; de Vargas et al., and fungi immediately come to mind when we hear 2015). In particular, several prevailing and very abun- the term “eukaryotes”. However, these groups of dant ribogroups such as MALV, MAST, MAOP, organisms represent a minority in the real diversity MAFO (marine alveolates, stramenopiles, opistho- of evolutionary lineages of eukaryotes. -

The Apicoplast: a Review of the Derived Plastid of Apicomplexan Parasites
Curr. Issues Mol. Biol. 7: 57-80. Online journalThe Apicoplastat www.cimb.org 57 The Apicoplast: A Review of the Derived Plastid of Apicomplexan Parasites Ross F. Waller1 and Geoffrey I. McFadden2,* way to apicoplast discovery with studies of extra- chromosomal DNAs recovered from isopycnic density 1Botany, University of British Columbia, 3529-6270 gradient fractionation of total Plasmodium DNA. This University Boulevard, Vancouver, BC, V6T 1Z4, Canada group recovered two DNA forms; one a 6kb tandemly 2Plant Cell Biology Research Centre, Botany, University repeated element that was later identifed as the of Melbourne, 3010, Australia mitochondrial genome, and a second, 35kb circle that was supposed to represent the DNA circles previously observed by microscopists (Wilson et al., 1996b; Wilson Abstract and Williamson, 1997). This molecule was also thought The apicoplast is a plastid organelle, homologous to to be mitochondrial DNA, and early sequence data of chloroplasts of plants, that is found in apicomplexan eubacterial-like rRNA genes supported this organellar parasites such as the causative agents of Malaria conclusion. However, as the sequencing effort continued Plasmodium spp. It occurs throughout the Apicomplexa a new conclusion, that was originally embraced with and is an ancient feature of this group acquired by the some awkwardness (“Have malaria parasites three process of endosymbiosis. Like plant chloroplasts, genomes?”, Wilson et al., 1991), began to emerge. apicoplasts are semi-autonomous with their own genome Gradually, evermore convincing character traits of a and expression machinery. In addition, apicoplasts import plastid genome were uncovered, and strong parallels numerous proteins encoded by nuclear genes. These with plastid genomes from non-photosynthetic plants nuclear genes largely derive from the endosymbiont (Epifagus virginiana) and algae (Astasia longa) became through a process of intracellular gene relocation. -

Transfer of Nuclei Froma Parasite to Its Host
Proc. Natl. Acad. Sci. USA Vol. 81, pp. 5420-5424, September 1984 Botany Transfer of nuclei from a parasite to its host (Polysiphonia/Choreocolax/microspectrofluorometry) LYNDA J. GOFF* AND ANNETTE W. COLEMANt *Center for Coastal Marine Science and Department of Biology, University of California, Santa Cruz, CA 95064; and tDivision of Biology and Medicine, Brown University, Providence, RI 02912 Communicated by Kenneth V. Thimann, April 27, 1984 ABSTRACT During the normal course of infection, nuclei are transferred via secondary pit connections from the parasit- ic marine red alga Choreocolax to its red algal host Polysi- phonia. These "planetic" nuclei are transmitted by being cut off into specialized cells (conjunctor cells) that fuse with an adjacent host cell, thereby delivering parasite nuclei and other cytoplasmic organelles into host cell cytoplasm. Within the for- eign cytoplasm, planetic nuclei survive for several weeks and may be active in directing the host cellular responses to infec- tion, since these responses are seen only in host cells containing planetic nuclei. The transfer and long-term survival ofa nucle- us from one genus into the cytoplasm of another through mechanisms that have evolved in nature challenge our under- standing of nuclear-cytoplasmic interactions and our concept of "individual." Parasitic organisms have evolved many specialized mecha- nisms for invading their hosts. However, no example yet has been reported of the regular introduction of nuclei of a para- site into cytoplasm of living cells of its host, leading to modi- fication of the metabolism of the host cell to the benefit of the parasite. Such an interaction would presumably require a most intimate coordination of host and parasite metabolism. -

An Ultrastructural Study of Vegetative Cells of the Chromophyte
MORPHOLOGICAL & PHYLOGENETIC ANALYSIS OF TWO SPECIES OF HETEROKONT ALAGE Ian Misner A Thesis Submitted to the University of North Carolina at Wilmington in Partial Fulfillment Of the Requirements for the Degree of Master of Science Department of Biological Sciences University of North Carolina at Wilmington 2004 Approved by Advisory Committee _______________________________ ______________________________ Chair _______________________________ ______________________________ Accepted by ______________________________ Dean, Graduate School TABLE OF CONTENTS ABSTRACT ...................................................................................................................... iii ACKNOWLEDGMENTS ................................................................................................. v DEDICATION .................................................................................................................. vi LIST OF TABLES ............................................................................................................ vii LIST OF FIGURES ........................................................................................................... viii CHAPTER 1. PHYLOGENETIC POSITIONS OF THE COLORLESS, COLONIAL IRON-FLAGELLATE ANTHOPHYSA VEGETANS AND POLYKARYON PYRENOIDOSUM GEN. ET COMB. NOV. (HETEROKONTOPHYTA) .............. 1 Introduction ............................................................................................................ 1 Materials and Methods ........................................................................................... -

758 the Ultrastructure of an Alloparasitic Red Alga Choreocolax
PHYCOLOGIA 12(3/4) 1973 The ultrastructure of an alloparasitic red alga Choreocolax polysiphoniae I PAUL KUGRENS Department of Botany and Plant Pathology, Colorado State University, Fort Collins, Colorado 80521, U.S.A. AND JOHN A. WEST Department of Botany, University of California, Berkeley, California 94720, U.S.A. Accepted June 18, 1973 An alloparasite, Choreocolax polysipiloniae, apparently represents one of the most evolved parasitic red algae. Chlo�oplasts are highly redu�ed and consist of dOl!ble membrane limited organelles lacking any inter nal thylako!� developmen!. The unInucleate cells have thick walls, an absence of starch in cortical cells and larg� quantIties of starch In meduII ary cells. Host-para�ite connections are made by typical red algal pit con . nectIOns. G.eneral effects of t�e InfectIOn on the host .Include cell hypertrophy, decrease in floridean starch granules, dispersed cytoplasmiC matrIces, and contorsJOn of chloroplasts. Phycologia, 12(3/4): 175-186, 1973 Introduction of the host, Cryptopleura. Her decision was The paraSItIc red algae constitute a unique based on the similarity in reproductive struc 1?irou of organisms about which surprisingly tures between the host and parasite, and she � suggested bacteria as causal agents for such lIttle IS known, although their distinctive nature . has been recognized since the late nineteenth proliferatIons. Chemin (1937) also indicated century. There are approximately 40 genera, that bacteria might be causal agents since bac unknown numbers of species, and all are ex teria were isolated from surface-sterilized thalli clusively florideophycean, belonging to all of Callocolax neglectus. Observations on Lobo orders except the Nemaliales. -

Evidence for Endosymbiotic Gene Transfer and the Early Evolution of Photosynthesis
Evolution of Glutamine Synthetase in Heterokonts: Evidence for Endosymbiotic Gene Transfer and the Early Evolution of Photosynthesis Deborah L. Robertson and Aure´lien Tartar Biology Department, Clark University Although the endosymbiotic evolution of chloroplasts through primary and secondary associations is well established, the evolutionary timing and stability of the secondary endosymbiotic events is less well resolved. Heterokonts include both photosynthetic and nonphotosynthetic members and the nonphotosynthetic lineages branch basally in phylogenetic reconstructions. Molecular and morphological data indicate that heterokont chloroplasts evolved via a secondary endo- symbiosis, involving a heterotrophic host cell and a photosynthetic ancestor of the red algae and this endosymbiotic event may have preceded the divergence of heterokonts and alveolates. If photosynthesis evolved early in this lineage, nuclear genomes of the nonphotosynthetic groups may contain genes that are not essential to photosynthesis but were derived from the endosymbiont genome through gene transfer. These genes offer the potential to trace the evolutionary history of chloroplast gains and losses within these lineages. Glutamine synthetase (GS) is essential for ammonium assimilation and glutamine biosynthesis in all organisms. Three paralogous gene families (GSI, GSII, and GSIII) have been identified and are broadly distributed among prokaryotic and eukaryotic lineages. In diatoms (Heterokonta), the nuclear-encoded chloroplast and cytosolic-localized GS isoforms are encoded by members of the GSII and GSIII family, respectively. Here, we explore the evolutionary history of GSII in both photosynthetic and nonphotosynthetic heterokonts, red algae, and other eukaryotes. GSII cDNA sequences were obtained from two species of oomycetes by polymerase chain reaction amplification. Additional GSII sequences from eukaryotes and bacteria were obtained from publicly available databases and genome projects. -
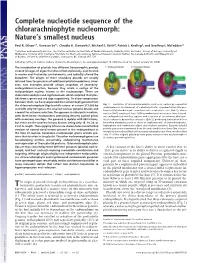
Complete Nucleotide Sequence of the Chlorarachniophyte Nucleomorph: Nature’S Smallest Nucleus
Complete nucleotide sequence of the chlorarachniophyte nucleomorph: Nature’s smallest nucleus Paul R. Gilson*†, Vanessa Su†‡, Claudio H. Slamovits§, Michael E. Reith¶, Patrick J. Keeling§, and Geoffrey I. McFadden‡ʈ *Infection and Immunity Division, The Walter and Eliza Hall Institute of Medical Research, Parkville 3050, Australia; ‡School of Botany, University of Melbourne, Victoria 3010, Australia; ¶Institute for Marine Biosciences, National Research Council, Halifax, NS, Canada B3H 3Z1; and §Department of Botany, University of British Columbia, Vancouver, BC, Canada V6T 1Z4 Edited by Jeffrey D. Palmer, Indiana University, Bloomington, IN, and approved April 19, 2006 (received for review January 26, 2006) The introduction of plastids into different heterotrophic protists created lineages of algae that diversified explosively, proliferated in marine and freshwater environments, and radically altered the biosphere. The origins of these secondary plastids are usually inferred from the presence of additional plastid membranes. How- ever, two examples provide unique snapshots of secondary- endosymbiosis-in-action, because they retain a vestige of the endosymbiont nucleus known as the nucleomorph. These are chlorarachniophytes and cryptomonads, which acquired their plas- tids from a green and red alga respectively. To allow comparisons between them, we have sequenced the nucleomorph genome from the chlorarachniophyte Bigelowiella natans: at a mere 373,000 bp Fig. 1. Evolution of chlorarachniophytes such as B. natans by sequential endosymbioses. Enslavement of a photosynthetic, cyanobacterium-like pro- and with only 331 genes, the smallest nuclear genome known and karyote (Cb) introduces photosynthesis into a eukaryotic host (Euk 1), whose a model for extreme reduction. The genome is eukaryotic in nature, nucleus (Nu1) acquires at least 1,000 cyanobacterial genes over time. -

Seven Gene Phylogeny of Heterokonts
ARTICLE IN PRESS Protist, Vol. 160, 191—204, May 2009 http://www.elsevier.de/protis Published online date 9 February 2009 ORIGINAL PAPER Seven Gene Phylogeny of Heterokonts Ingvild Riisberga,d,1, Russell J.S. Orrb,d,1, Ragnhild Klugeb,c,2, Kamran Shalchian-Tabrizid, Holly A. Bowerse, Vishwanath Patilb,c, Bente Edvardsena,d, and Kjetill S. Jakobsenb,d,3 aMarine Biology, Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316 Oslo, Norway bCentre for Ecological and Evolutionary Synthesis (CEES),Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316 Oslo, Norway cDepartment of Plant and Environmental Sciences, P.O. Box 5003, The Norwegian University of Life Sciences, N-1432, A˚ s, Norway dMicrobial Evolution Research Group (MERG), Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316, Oslo, Norway eCenter of Marine Biotechnology, 701 East Pratt Street, Baltimore, MD 21202, USA Submitted May 23, 2008; Accepted November 15, 2008 Monitoring Editor: Mitchell L. Sogin Nucleotide ssu and lsu rDNA sequences of all major lineages of autotrophic (Ochrophyta) and heterotrophic (Bigyra and Pseudofungi) heterokonts were combined with amino acid sequences from four protein-coding genes (actin, b-tubulin, cox1 and hsp90) in a multigene approach for resolving the relationship between heterokont lineages. Applying these multigene data in Bayesian and maximum likelihood analyses improved the heterokont tree compared to previous rDNA analyses by placing all plastid-lacking heterotrophic heterokonts sister to Ochrophyta with robust support, and divided the heterotrophic heterokonts into the previously recognized phyla, Bigyra and Pseudofungi. Our trees identified the heterotrophic heterokonts Bicosoecida, Blastocystis and Labyrinthulida (Bigyra) as the earliest diverging lineages. -

Organellar Genome Evolution in Red Algal Parasites: Differences in Adelpho- and Alloparasites
University of Rhode Island DigitalCommons@URI Open Access Dissertations 2017 Organellar Genome Evolution in Red Algal Parasites: Differences in Adelpho- and Alloparasites Eric Salomaki University of Rhode Island, [email protected] Follow this and additional works at: https://digitalcommons.uri.edu/oa_diss Recommended Citation Salomaki, Eric, "Organellar Genome Evolution in Red Algal Parasites: Differences in Adelpho- and Alloparasites" (2017). Open Access Dissertations. Paper 614. https://digitalcommons.uri.edu/oa_diss/614 This Dissertation is brought to you for free and open access by DigitalCommons@URI. It has been accepted for inclusion in Open Access Dissertations by an authorized administrator of DigitalCommons@URI. For more information, please contact [email protected]. ORGANELLAR GENOME EVOLUTION IN RED ALGAL PARASITES: DIFFERENCES IN ADELPHO- AND ALLOPARASITES BY ERIC SALOMAKI A DISSERTATION SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOLOGICAL SCIENCES UNIVERSITY OF RHODE ISLAND 2017 DOCTOR OF PHILOSOPHY DISSERTATION OF ERIC SALOMAKI APPROVED: Dissertation Committee: Major Professor Christopher E. Lane Jason Kolbe Tatiana Rynearson Nasser H. Zawia DEAN OF THE GRADUATE SCHOOL UNIVERSITY OF RHODE ISLAND 2017 ABSTRACT Parasitism is a common life strategy throughout the eukaryotic tree of life. Many devastating human pathogens, including the causative agents of malaria and toxoplasmosis, have evolved from a photosynthetic ancestor. However, how an organism transitions from a photosynthetic to a parasitic life history strategy remains mostly unknown. Parasites have independently evolved dozens of times throughout the Florideophyceae (Rhodophyta), and often infect close relatives. This framework enables direct comparisons between autotrophs and parasites to investigate the early stages of parasite evolution.